三葉蟲
| 三叶虫纲 化石时期:530–251.4 Ma PreЄ Є O S D C P T J K Pg N | |
|---|---|
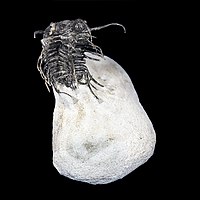 | |
Koneprusia brutoni | |
科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 节肢动物门 Arthropoda |
| 演化支: | 蛛形类 Arachnomorpha |
| 亚门: | †三叶虫亚门 Trilobitomorpha |
| 纲: | †三叶虫纲 Trilobita Walch, 1771 |
目 | |
| |
三叶虫纲(學名:Trilobita)的动物通称三叶虫,是节肢动物门中已经灭绝的一纲。牠们最早出现于寒武紀,在古生代早期达到顶峰,此后逐渐减少至灭绝。最晚的三叶虫(砑头虫目)于二亿五千万年前二叠纪结束时的生物集群灭绝中消失。三叶虫是非常知名的化石动物,其知名度可能仅次于恐龙。在所有的化石动物中三叶虫是种类最丰富的,至今已经确定的有九(或者十)个目,一万五千多个物种。大多数三叶虫是比较简单的、小的海生动物,牠们在海底爬行,通过过滤泥沙来吸取营养。牠们身體分節,有帶溝將身體分為三個垂直的葉。在世界各地都有發現過其化石。
目录
1 形态
2 感觉器官
3 发育
4 术语
5 来源
6 灭绝
7 化石分布
8 分類
9 参考资料
10 外部链接
形态
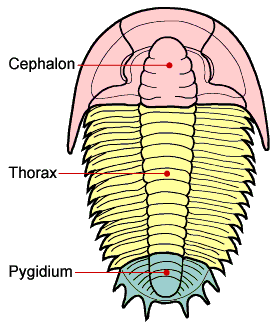
三叶虫的躯体分三个体段(tagmata):头部由口前的两个环和口后的四个环完全融合在一起组成,胸部由可以相互运动的环组成,尾部由最后几个羽尾扇和完全融合在一起的环组成。最原始的三叶虫尾部的環構造相当简单。三叶虫的胸部非常灵活——化石的三叶虫有時會像現今的地鳖一样可以將身體卷在一起来保护自己。
三叶虫有一对口前的触角,牠的其他足之间没有区别。每个足有六个节,这与其他早期的节肢动物类似。第一节还带有羽毛似的副叶被用来呼吸和游泳。躯体上有从中叶伸出的侧叶。这个横向的三叶结构是三叶虫名字的来源,而不是牠纵向分为头、胸、尾三部分。
虽然三叶虫只在背部有盔甲,但是牠们的外骨骼还是相当重的,牠们的外骨骼是由甲壳素为主的蛋白质联合方解石和磷化钙等矿物组成的。不像其他节肢动物那样能够在蜕皮前重新吸收外骨骼中的大部分矿物,三叶虫在蜕皮是将所有盔甲中的矿物全部抛弃,因此一个三叶虫可以留下多个良好地矿物化的外骨骼,这提高了三叶虫化石的数量。在蜕皮时外骨骼首先在头部和胸部之间分开,这是为什么许多三叶虫的化石不是缺少头部就是缺少胸部,其实许多化石是三叶虫蜕掉的皮,而不是死去的三叶虫形成的。大多数三叶虫的头部有两个面部缝合来简化蜕皮过程。头部的两侧有一对复眼,有些种的复眼相当先进。事实上约5.43亿年前三叶虫是第一批进化出真正的眼睛的动物。有人认为眼睛的出现是寒武纪生命大爆发的导致原因。
从奥陶纪到泥盆纪末,一些三叶虫(比如裂肋三叶虫目)进化出了非常巧妙的棘刺(spines)似的结构,尤其在摩洛哥发现了这样的化石。不过要当心的是许多从摩洛哥出售的带有棘刺结构的三叶虫化石实际上是伪造品。此外在俄罗斯西部、美国俄克拉何马州以及加拿大安大略省也有带棘刺结构的化石被发现。这种棘刺结构可能是对于鱼的出现的一种抵抗反应。
据《新科学家》(2005年5月)的报道“有些……三叶蟲……的头上有类似现代甲虫的角。”根据这些角的形状、大小和位置伦敦大学玛利皇后学院的罗布·克奈尔(Rob Knell)和伦敦自然历史博物馆的理查德·福提得出结论认为它们用来作为寻找配偶时进行角斗。假如这个理论正确的话,三叶虫是进化史上最早表现这个行为的动物。
三叶虫的大小在1毫米至90厘米之间,典型的大小在2至7厘米间。最大的三叶虫是2009年在葡萄牙阿洛卡地质公园里奥陶纪的岩石里被发现的。
感觉器官


蛙形镜眼虫的复眼

复眼内透镜数較多之拟平鬃虫

手尾蟲(Cheirurus sp.),产于摩洛哥,泥盆纪

碩寬展櫛蟲(Asaphus expansus robustus),产于俄罗斯,奥陶纪
许多三叶虫有眼睛,牠们还有可能用来作味觉和嗅觉器官的触角。有些三叶虫是瞎的,可能牠们居住在非常深的海底,那里没有光,因此用不着眼睛。有些(比如蛙形镜眼虫Phacops rana)有很大的眼睛。
三叶虫的眼睛是由方解石(碳酸钙,CaCO3)组成的。纯的方解石是透明的,有些三叶虫使用单晶的、透明的方解石来组成其每只眼睛的透镜。这与大多数其他节肢动物不同,大多数节肢动物使用软透镜的、由甲壳素组成的眼睛。三叶虫坚固的方解石透镜无法像人的软晶状体的眼睛那样来调节焦距。但是有些三叶虫的方解石组成一个内部的、复合结构,这个结构可以降低球差,同时提供极好的景深。在今天生存的动物中蛇尾海星Ophiocoma wendtii使用类似的透镜。
典型的三叶虫眼睛是复眼,每个透镜都是一个拉长的棱镜。每只复眼内的透镜数不等,有些只有一个,有些可达上千。在这样的复眼中其透镜一般排列为六边形。三叶虫的眼睛可分為全色眼和裂色眼兩種。
发育
从卵中孵化出来的幼虫被称为原甲期(protaspid),在这个阶段里所有环全部融合在一起形成一个单一的盔甲。在此后的生长期里在每次蜕皮时在尾部前会增加新的胸部环。此后在蜕皮时环的数目不再增加。对三叶虫的幼虫阶段人们的认识很丰富,牠们为研究三叶虫之间的亲缘关系提供了非常重要的帮助。
术语
在描述不同属的区别时,头部不同部位的存在、大小和形状特别重要。
图1显示头部的大约形态。面部是头鞍(glabella)两边的侧叶。三叶虫蜕皮或者死后其活動頰(librigena)往往会分开,暴露其头盖(cranidium),图2显示了头部更细致的结构。
 图1—头部的结构 |  图2—头部的细致结构 |
来源

鬼鞍角蟲的一種(Cyphaspis tafilalet),产于摩洛哥,泥盆纪

越南溝通蟲(Ductina vietnamica),产于中国,泥盆纪
基于形态上的类似三叶虫的祖先可能是类似于节肢动物的动物如斯普里格蠕虫或其他隐生宙埃迪卡拉纪时期类似三叶虫的动物。早期三叶虫与伯吉斯页岩和其他寒武纪的节肢动物化石有许多类似的地方[1]。因此三叶虫与其他节肢动物可能在埃迪卡拉纪和寒武纪的交界之前有共同的祖先。
灭绝

卡瓦勒斯基櫛蟲(Asaphus kowalewskii),一种在俄罗斯出土的三叶虫

二叶石,三叶虫足迹的化石
三叶虫灭绝的具体原因不明,但是志留紀和泥盆纪时期两顎有強大关节連接的鲨鱼和其他早期鱼类的出现,与同时发生的三叶虫数量的减少似乎不是无关的。三叶虫可能为这些新动物提供了丰富的食物。
此外到二叠纪后期时三叶虫的数量和种类已经相当少了,僅剩下砑頭蟲目(Proetida),这无疑为牠们在二叠纪-三叠纪灭绝事件中灭绝提供了条件。此前的奥陶纪-志留纪灭绝事件虽然没有后来的二叠纪-三叠纪灭绝事件那么严重,但是也已经大大地减少了三叶虫的多样性。
而晚寒武世海洋深部缺氧也是直接导致三叶虫灭绝的原因[2]。
今天存在与三叶虫最接近的动物可能是鲎或头虾纲(Cephalocarida)等动物。
化石分布
清初文人王士禎在《池北偶谈》一書《谈艺》篇中首次记载了泰山燕子石,即三叶虫化石,“背负一小蝠、一蚕,腹下蝠近百,飞者、伏者、肉羽如生,蚕右天然有小凹,可以受水;下方正,受墨。公制为砚,名曰:‘多福砚’”。由于三叶虫总是与其他海洋动物的化石一起被发现,因此它们看来全部在海洋中生活。在远古海洋中三叶虫的生活环境从浅海到深海非常广。偶尔三叶虫在海底爬行时留下的足迹也被化石化了。几乎在所有今天的大陆上均有三叶虫的化石被发现,牠们似乎在所有远古海洋中均有生存。
今天在全世界发现的三叶虫化石可以分上万种,由于三叶虫的发展非常快,因此牠们非常适合被用作标准化石,地质学家可以使用牠们来确定含有三叶虫的石头的年代。三叶虫是最早的、获得广泛吸引力的化石,至今为止每年还有新的物种被发现。一些印第安人部落认识到三叶虫是水生动物,他们称三叶虫为“石头里的小水虫”。
三葉蟲是美國俄亥俄州(Isotelus)、威斯康辛州(Calymene)和宾夕法尼亚州(Phacops rana)的州化石。
在英属哥伦比亚、纽约州、中国、德国和其他一些地方发现过非常稀有的、带有软的身体部位如足、鳃和触角的三叶虫化石。
在俄罗斯、德国、摩洛哥、美国和加拿大均有商业采集三叶虫化石的企业。
2000年於加拿大發現最大化石三葉蟲Isotelus rex,長720毫米,寬400毫米,時間約4億4千5百萬年前奧陶紀時期的。
分類

三葉蟲綱各目的演化圖
萊得利基蟲目(Redlichiida)(早寒武紀~中寒武紀):為最原始的三葉蟲,被認為是三葉蟲綱其他目的祖先,也是目前已知最早出現在化石上的節肢動物。擁有細小的尾巴和較扁平的身軀,以及巨大的半月形全色眼。
球接子目(Agnostida)(早寒武紀~晚奧陶紀):身體較小,尾部與頭部的形狀大小相等,大部分沒有眼睛。可能後來演化成甲殼類。
褶頰蟲目(Ptychopariida)(早寒武紀~晚奧陶紀):由萊德利基蟲目演化而成,包括的三葉蟲種類相當的繁多,其後再演化成其他三葉蟲目。擁有較大的胸部,通常比尾部還要長。
櫛蟲目(Asaphida)(早寒武紀~晚志留紀):由褶頰蟲目演化而成。體型較大,頭部與尾部通常大小相近,多數品種外型扁平,具有強壯的尾部。
縱棒頭蟲目(Corynexochida)(早寒武紀~晚泥盆紀):由褶頰蟲目演化而成,是分佈範圍極其廣泛的三葉蟲類別,各大洲都有其代表。
裂肋蟲目(Lichida)(中寒武紀~中泥盆紀):這類三葉蟲未有確實的祖先資料。身軀長滿小刺。小刺主要用作保護身體,免受襲擊,而且具有平衡的作用。有人將裂肋蟲目齒肋蟲科(Odontopleuridae)和德氏蟲科(Damesellidae)獨立出來,變成齒肋蟲目(Odontopleurida)。
鏡眼蟲目(Phacopida)(早奧陶紀~晚泥盆紀):這類三葉蟲未有確實的祖先資料。具有發達的複眼──裂色眼,對環境有很強的適應能力,多具有發達的捲曲器官。此類群的滅絕標誌著三葉蟲繁盛海洋時代的終結。
砑頭蟲目(Proetida)(中寒武紀~晚二疊紀):泥盆紀後期的大滅絕中唯一倖存的三葉蟲類,具有流線型的身體,但身上沒有刺和裝飾。
鐮蟲目(Harpetida)(中寒武紀~晚泥盆紀):這類三葉蟲無吻板(rostral plate),且面縫合線(facial suture)的樣式十分特殊,是一種沿著頭部外緣的縫合線型式。
游盾目(Nektaspida)(早寒武紀~晚志留紀):包括納羅蟲、周小姐蟲等。
维基共享资源中相关的多媒体资源:三葉蟲 |
维基物种中的分类信息:三葉蟲 |
参考资料
^ http://www.trilobites.info/triloclass.htm
^ Li, Dandan; Zhang, Xiaolin; Hu, Dongping; Chen, Xiaoyan; Huang, Wei; Zhang, Xu; Li, Menghan; Qin, Liping; Peng, Shanchi; Shen, Yanan. Evidence of a large δ13Ccarb and δ13Corg depth gradient for deep-water anoxia during the late Cambrian SPICE event. Geology. 2018-06-12, 46 (7): 631–634. doi:10.1130/G40231.1.
外部链接
- 吉林大学博物馆
- 石尚小學堂
|


