病毒
| 病毒界 | |
|---|---|
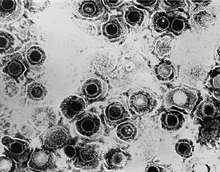 | |
| 簡單的疱疹病毒1(HSV-1) | |
科学分类 | |
| 界: | 病毒界 Virus |
| 组别 | |
| |
一個位於宿主細胞之外的獨立、功能完全的病毒顆粒
在病毒基因外包裹着的一層蛋白質外殼。衣殼由許多相同的、分子較小的蛋白質(這些蛋白質稱爲「殼粒」(capsomer))組成
病毒包膜(包膜)
一些病毒擁有的包裹病毒體的脂肪泡
一段DNA或RNA。如果把核苷酸比作字的話,那麼基因就是由核苷酸寫成的句子。基因會指導病毒的複製。一種病毒只能擁有DNA與RNA中的一種作爲它的遺傳物質。
能被一種或多種病毒感染的動物、植物,或者細菌
噬菌體
能在細菌體內繁殖的病毒
細胞嗜性決定了會被病毒選爲繁殖場所的細胞的類型
病毒表面的抗原不同,血清型也就不同
對稱性
一種病毒如果(不同個體間)完全相同,而且具有立方體、螺旋形等複雜的結構,那麼它就具有對稱性。
病毒(英语:virus,中文舊稱“濾過性病毒”)是由一个核酸分子(DNA或RNA)与蛋白质构成的非细胞形态,靠寄生生活的介於生命体及非生命體之間的有機物種,它既不是生物亦不是非生物,目前不把它歸於五界(原核生物、原生生物、真菌、植物和動物)之中。[1]它是由一个保护性外壳包裹的一段DNA或者RNA,藉由感染的機制,这些简单的有機体可以利用宿主的细胞系统进行自我复制,[2]但无法独立生长和复制。病毒可以感染几乎所有具有细胞结构的生命体。第一个已知的病毒是烟草花叶病毒,由马丁乌斯·贝杰林克于1899年发现并命名,[3]迄今已有超过5000种类型的病毒得到鉴定。[4]研究病毒的科学称为病毒学,是微生物学的一个分支。
病毒由两到三个成份组成:病毒都含有遺傳物質(RNA或DNA,只由蛋白质组成的朊毒體并不属于病毒[5]);所有的病毒也都有由蛋白质形成的衣壳,用来包裹和保护其中的遗传物质;此外,部分病毒在到达细胞表面时能够形成脂质包膜环绕在外。病毒的形态各异,从简单的螺旋形和正二十面體形到複合型结构。病毒颗粒大约是细菌大小的百分之一。[6]病毒的起源目前尚不清楚,不同的病毒可能起源于不同的机制:部分病毒可能起源于质粒(一种环状的DNA,可以在细胞内复制并在细胞间进行转移),而其他一些则可能起源于细菌。
病毒的传播方式多种多样,不同类型的病毒采用不同的方法。例如,植物病毒可以通过以植物汁液为生的昆虫,如蚜虫,来在植物间进行传播;而动物病毒可以通过蚊虫叮咬而得以传播。这些携带病毒的生物体称为“载体”。流感病毒可以经由咳嗽和打喷嚏来传播;诺罗病毒则可以通过手足口途径来传播,即通过接触带有病毒的手、食物和水;輪狀病毒常常是通过接触受感染的儿童而直接传播的;此外,艾滋病毒则可以通过性接触来传播。
并非所有的病毒都会导致疾病,因为许多病毒的复制并不会对受感染的器官产生明显的伤害。一些病毒,如艾滋病毒,可以与人体长时间共存,并且依然能保持感染性而不受到宿主免疫系统的影响,即“病毒持续感染”(viral persistence)。[7]但在通常情况下,病毒感染能够引发免疫反应,消灭入侵的病毒。而这些免疫反应能够通过注射疫苗来产生,从而使接种疫苗的人或动物能够终生对相应的病毒免疫。像细菌这样的微生物也具有抵御病毒感染的机制,如限制修飾系統。[8]抗生素对病毒没有任何作用,但用于治疗病毒感染的抗病毒药物已經研发出來。[9]
目录
1 歷史
2 起源
3 结构
3.1 螺旋形
3.2 正二十面體形
3.3 包膜型
3.4 複合型
4 遗传物质
4.1 遗传变异
5 生命周期
5.1 DNA病毒
5.2 RNA病毒
5.3 反转录病毒
6 命名
7 分類
7.1 ICTV分类法
7.2 巴尔的摩分类法
8 病毒與人類疾病
8.1 流行病学
8.2 流行病和瘟疫
8.3 癌症
8.4 宿主防御机制
8.5 预防与治疗
8.5.1 疫苗
8.5.2 抗病毒药物
9 其他物种中的病毒感染
9.1 动物
9.2 植物
9.3 细菌
9.4 古菌
10 应用
10.1 生命科学与医学
10.2 材料科学与纳米技术
10.3 武器
11 参见
12 參考文獻
12.1 引用
12.2 书籍
13 延伸阅读
14 外部链接
歷史

荷兰微生物学家马丁乌斯·贝杰林克
关于病毒所导致的疾病,早在公元前2-3世纪的印度和中国就有了关于天花的記錄。[10]但直到19世纪末,病毒才开始逐渐得以发现和鉴定。1884年,法国微生物学家查理斯·尚柏朗发明了一种细菌无法滤过的过滤器(尚柏朗過濾器,其滤孔孔径小于细菌的大小),他利用这一过滤器就可以将液体中存在的细菌除去。[11] 1892年,俄国生物学家德米特里·伊凡諾夫斯基在研究烟草花叶病时发现,将感染了花叶病的烟草叶的提取液用烛形滤器过滤后,依然能够感染其他烟草。于是他提出这种感染性物质可能是细菌所分泌的一种毒素,但他并未深入研究下去。[12]当时,人们认为所有的感染性物质都能够藉由过滤除去并且能够在培养基中生长,这也是疾病的细菌理论(germ theory)的一部分。[13] 1899年,荷兰微生物学家马丁乌斯·贝杰林克重复了伊凡諾夫斯基的实验,并相信这是一种新的感染性物质。[14]他还观察到这种病原只在分裂细胞中复制,由于他的实验没有显示这种病原的颗粒形态,因此他称之为contagium vivum fluidum(可溶的活菌)并进一步命名为virus(病毒)。[12]贝杰林克认为病毒是以液态形式存在的(但这一看法后来由温德尔·梅雷迪思·斯坦利推翻,他证明了病毒是颗粒状的)。[12]同样在1899年,弗雷德里希·勒夫勒(Friedrich Loeffler)和保羅·弗羅施(Paul Frosch)发现患口蹄疫动物淋巴液中含有能通过滤器的感染性物质,由于经过了高度的稀释,排除了其为毒素的可能性;他们推论这种感染性物质能够自我复制。[12]
20世纪早期,英国细菌学家弗雷德里克·圖爾特发现了可以感染细菌的病毒,并称之为噬菌体。[15]随后法裔加拿大微生物学家費里斯·代列爾(Félix d'Herelle)描述了噬菌体的特性:将其加入长满细菌的琼脂固体培养基上,一段时间后会出现由于细菌死亡而留下的空斑。高浓度的病毒悬液会使培养基上的细菌全部死亡,但通过精确的稀释,可以产生可辨认的空斑。通过计算空斑的数量,再乘以稀释倍数就可以得出溶液中病毒的个数。[16]他们的工作揭开了现代病毒学研究的序幕。
在19世纪末,认为病毒的特性是感染性、可滤过性和需要活的宿主,也就意味着病毒只能在动物或植物体内生长。1906年,哈里森发明了在淋巴液中进行组织生长的方法;接着在1913年,E·斯坦哈特(E. Steinhardt)、C·伊斯雷利(C. Israeli)和R·A·蘭伯特(R. A. Lambert)利用这一方法在豚鼠角膜组织中成功培养了牛痘苗病毒,突破了病毒需要体内生长的限制。
[17] 1928年,H·B·梅特蘭(H. B. Maitland)和M·C·梅特蘭(M. C. Maitland)有了更进一步的突破,他们利用切碎的母鸡肾臟的悬液对牛痘苗病毒进行了培养。他们的方法在1950年代得以广泛应用于脊髓灰質炎病毒疫苗的大规模生产。[18]
另一项研究突破发生在1931年,美國病理学家歐內斯特·古巴斯捷(Ernest William Goodpasture)在受精的鸡蛋中培养了流感病毒。[19] 1949年,约翰·富兰克林·恩德斯、托马斯·哈克尔·韦勒和弗雷德里克·查普曼·罗宾斯利用人的胚胎细胞对脊髓灰質炎病毒进行了培养,这是首次在没有固体动物组织或卵的情况下对病毒进行的成功培养。乔纳斯·索尔克利用此一研究成果来有效生产脊髓灰質炎病毒疫苗。[20]
1931年,德国工程师恩斯特·鲁斯卡和马克斯·克诺尔发明了电子显微镜,使得研究者首次得到了病毒形态的照片。[21] 1935年,美国生物化学家和病毒学家温德尔·梅雷迪思·斯坦利发现烟草花叶病毒大部分是由蛋白质所组成的,并得到病毒晶体。[22]随后,他将病毒成功地分离为蛋白质部分和RNA部分。[23]温德尔·斯坦利也因为他的这些发现而获得了1946年的诺贝尔化学奖。[24]烟草花叶病毒是第一个结晶的病毒,从而可以通过X射线晶体学的方法来得到其结构细节。第一张病毒的X射线衍射照片是由貝納(Bernal)和范库肯(Fankuchen)于1941年所拍摄的。1955年,通过分析病毒的衍射照片,羅莎琳·富蘭克林揭示了病毒的整体结构。[25]同年,海因茨·弗羅倫克-卡納特(Heinz Fraenkel-Conrat)和羅布利·威廉姆斯发现将分离纯化的烟草花叶病毒RNA和衣壳蛋白混合在一起后,可以重新组装成具有感染性的病毒,这也揭示了这一简单的机制很可能就是病毒在它们的宿主细胞内的组装过程。[26]
20世纪的下半叶是发现病毒的黄金时代,大多数能够感染动物、植物或细菌的病毒在这数十年间发现。[3][27] 1957年,马动脉炎病毒和导致牛病毒性腹泻的病毒(一种瘟病毒)正式发现;1963年,巴鲁克·塞缪尔·布隆伯格发现了乙型肝炎病毒;[28] 1965年,霍华德·马丁·特明发现并描述了第一种逆转录病毒;这类病毒将RNA逆转录为DNA的关键酶,即逆转录酶,在1970年由霍华德·马丁·特明和戴维·巴尔的摩分别独立鉴定出来。[29] 1983年,法国巴斯德研究院的吕克·蒙塔尼耶和他的同事弗朗索瓦丝·巴尔-西诺西首次分离得到了一种逆转录病毒,也就是现在世人皆知的艾滋病毒(HIV)。[30]其二人也因此与发现了能够导致子宮頸癌的人乳头状瘤病毒的德国科学家哈拉尔德·楚尔·豪森分享了2008年的诺贝尔生理学与医学奖。[31]
起源
只要有生命的地方,就有病毒存在;病毒很可能在第一个细胞进化出来时就存在了。[32]病毒起源于何时尚不清楚,因为病毒不形成化石,也就没有外部参照物来研究其进化过程,同时病毒的多样性显示它们的进化很可能是多条线路的而非单一的。[33]分子生物学技术是目前可用的揭示病毒起源的方法;[34]但这些技术需要获得远古时期病毒DNA或RNA的样品,而目前储存在实验室中最早的病毒样品也不过90年。[35][36]有三种流行的关于病毒起源的理论:[37][38]
逆向理论(Regressive theory):病毒可能曾经是一些寄生在较大细胞内的小细胞。随着时间的推移,那些在寄生生活中非必需的基因逐渐丢失。这一理论的证据是,细菌中的立克次氏体和衣原體就像病毒一样,需要在宿主细胞内才能复制;而它们缺少了能够独立生活的基因,这很可能是由于寄生生活所导致的。这一理论又称为退化理论(degeneracy theory)。[39][40]
细胞起源理论(有时也称为漂荡理论):[39][41]一些病毒可能是从较大生物体的基因中“逃离”出来的DNA或RNA进化而来的。逃离的DNA可能来自质粒(可以在细胞间传递的裸露DNA分子)或转座子(可以在细胞基因内不同位置复制和移动的DNA片段,[42]曾称为“跳跃基因”,属于可移动遗传元件)。转座子是在1950年由巴巴拉·麦克林托克在玉米中发现的。[43]
共进化理论:病毒可能进化自蛋白质和核酸复合物,与细胞同时出现在远古地球,并且一直依赖细胞生命生存至今。类病毒是一类RNA分子,但不归入病毒中,因为它们缺少由蛋白质形成的衣壳。然而,它们具有多种病毒的普遍特征,常常称为亚病毒物质。[44]类病毒是重要的植物病原体。[45]它们没有编码蛋白质的基因,但可以与宿主细胞作用,利用宿主来进行它们自身的复制。[46]人類丁型肝炎病毒具有和类病毒相似的RNA基因组,也不能生成自己的蛋白质衣壳,但却能够利用乙型肝炎病毒的衣壳。因此,丁型肝炎病毒是一种缺陷型病毒,需要乙型肝炎病毒的帮助才能够进行复制。[47]这些依赖于其他种类病毒的病毒称为“卫星病毒”,它们可能是介于类病毒和病毒之间的进化中间体。[48][49]朊病毒是具有感染性的蛋白质分子,不含DNA或RNA。[50]朊病毒会导致绵羊感染羊搔痒症或牛感染牛海绵状脑病(俗稱“疯牛病”),也会使人获患庫魯病(Kuru)和克雅二氏病。[51]虽然缺乏核酸,朊病毒依然能够复制,这是因为在生物体内存在与朊病毒具有相同序列但结构不同的正常蛋白质,而朊病毒可以使这些正常蛋白质的结构发生变化,转化为朊病毒,这样新产生的朊病毒又可以感染更多的正常蛋白质,使得朊病毒越来越多。虽然朊病毒与病毒或类病毒本质完全不同,但朊病毒的发现进一步提高了该理论的可信性,说明病毒可能进化自能够自我复制的分子。[52]
利用计算机来分析病毒和宿主DNA的序列信息,可以对不同病毒之间的进化关系有更好的了解,而且可以有助于发现现代病毒的祖先。至今这类分析还没有能够决定哪一种理论是正确的。而且不大可能所有的病毒都来自同一祖先,不同的病毒很可能是通过一种或多种机制在不同的时期产生。[53]
对于病毒到底是一种生命形式,还是仅仅是一种能够与生物体作用的有机结构,人们的观点各不相同。有人将病毒描述为处于“生命边缘的生物体”,[54]因为它们像其它生物体一样拥有基因、能够通过自然选择而进化[55]并且能够通过自行组装来完成复制。然而,虽然病毒含有基因,但它们没有细胞结构,而细胞被认为是生命的基本单位;而且,病毒没有自身的代谢机制,需要通过宿主细胞来替它们完成复制繁殖,因此它们不能够在宿主细胞外进行繁殖(虽然属于细菌的立克次氏体和衣原體也具有同样的缺陷);可接受的生命形式是利用细胞分裂来进行繁殖的,而病毒是自发地在细胞内进行组装的,类似于晶体的自发生长过程。虽然病毒是否是一种生命形式,目前还没有定论,但病毒在宿主细胞内的自组装方式对于研究生命起源具有一定启示意义,[56]有一种假说就认为生命是起始于有机分子的自组装。[57]
结构

A.無包膜病毒;B.具包膜病毒。①衣殼,②核酸,③殼粒,④核衣殼,⑤病毒体,⑥外套膜,⑦刺突蛋白
病毒的形状和大小(统称形态)各异。大多数病毒的直径在10-300纳米(nm)。一些絲狀病毒的长度可达1400nm,但其宽度却只有约80nm。[6]大多数的病毒无法在光学显微镜下观察到,而扫描或透射电子显微镜是观察病毒颗粒形态的主要工具,常用的染色方法为负染色法。[58]
一个完整的病毒颗粒称为“病毒体”(virion),是由蛋白质组成的具有保护功能的“衣壳(Capsid,或稱蛋白質外殼)”和衣壳所包住的核酸组成。形成衣殼的等同的蛋白质亚基稱作次“蛋白衣”或“殼粒”(capsomere)。[59]有些病毒的核衣壳外面,还有一层由蛋白质、多糖和脂类构成的膜叫做“包膜”(envelope,又稱外套膜),包膜上生有“刺突”(spike),如流感病毒。衣壳是由病毒基因组所编码的蛋白质组成的,它的形状可以作为区分病毒形态的基础。[60][61]通常只需要存在病毒基因组,衣壳蛋白就可以自组装成为衣壳。但结构复杂的病毒还会编码一些帮助构建衣壳的蛋白质。与核酸结合的蛋白质称为核蛋白,核蛋白与核酸结合形成核糖核蛋白,再与衣壳蛋白结合在一起就形成了“核衣壳”。病毒的形态一般可以分为以下四种:
螺旋形

烟草花叶病毒结构:病毒RNA卷曲在的由重复的蛋白质亚基组成的螺旋形衣壳中。
螺旋形的衣壳是由壳粒绕着同一个中心轴排列堆积起来,以形成一个中空的棒状结构。这种棒状的病毒体可以是短而刚性的,也可以是长而柔性的。具有这种形态的病毒一般为单链RNA病毒,研究得最多的就是烟草花叶病毒,[62]但也有少量单链DNA病毒也为螺旋形;无论是哪一种病毒,其核酸都通过静电相互作用与衣壳蛋白结合(核酸带负电而衣壳蛋白朝向中心的部分带正电)。一般来说,棒状病毒体的长度取决于内部核酸的长度,而半径取决于壳粒的大小和排列方式。用于定义这种螺旋形态的参数有两个:amplitude和pitch,前者即直径,而后者是指壳粒环绕一周后所前进的距离。[63]
正二十面體形
具有正二十面体结构的腺病毒的电镜照片
大多数的动物病毒为正二十面体或具有正二十面体对称的近球形结构。二十面体具有5-3-2对称,即每个顶点为5重对称,每个面的中心为3重对称,每条边的中心为2重对称。[63]病毒之所以采用这种结构可能的一个很重要的原因是,规则的二十面体是相同壳粒形成封闭空间的一个最优途径,可以使所需的能量最小化。[64]形成二十面体所需的最少的等同的壳粒的数量为12,每个壳粒含有5个等同的亚基。但很少有病毒只含有60个衣壳蛋白亚基,多数正二十面体形病毒的亚基数量大于60,为60的倍数,倍数可以是3、4、7、9、12或更多。[64]由于二十面体的对称性,位于顶点的壳粒周围有五个壳粒环绕,称为“penton”;而位于三角形面中心的壳粒周围有六个壳粒环绕,称为“hexon”。[65]
包膜型
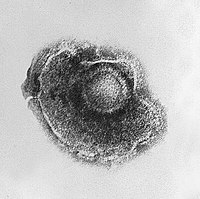
带状疱疹病毒的电镜负染照片显示其病毒颗粒周围環繞在包膜裡。
一些病毒可以利用改造后的宿主的细胞膜(来自细胞表面的质膜或细胞内部的膜,如核膜及内质网膜)环绕在病毒体周围,形成一层脂质的包膜。包膜上既镶嵌有来自宿主的膜蛋白也有来自病毒基因组编码的膜蛋白;而脂质膜本身和其中的糖类则都来自宿主细胞。包膜型病毒位于包膜内的病毒体可以是螺旋形或正二十面体形的。[63]
无包膜的病毒在宿主细胞内完成复制后,需要宿主细胞死亡并裂解后,才能逸出并进一步感染其他细胞。这种方法虽然简单,但常常造成大量非成熟细胞死亡,反而降低了对宿主细胞的利用率。而有了包膜之后,病毒可以通过包膜与宿主的细胞膜融合来出入细胞,而不需要造成细胞死亡。[63]流感病毒和艾滋病毒就采用的是这种策略。大多数的包膜型病毒的感染性都依赖于包膜。[66]
複合型

一个典型的有尾噬菌体的结构:①头部,②尾部,③核酸,④头殼,⑤颈部,⑥尾鞘,⑦尾丝,⑧尾钉,⑨基板
与以上三类病毒形态相比,复合型病毒的结构复杂得多,它们的衣壳既非完全的螺旋形又非完全的正二十面体形,可以有附加的结构,如蛋白质组成的尾巴或复杂的外壁。有尾噬菌体和痘病毒都是比较典型的复合型病毒。
有尾噬菌体在噬菌体中数量最多,其壳体由头部和尾部组成,头部呈正二十面体对称,尾部呈螺旋对称,头部和尾部之间通过颈部相连。此外噬菌体的尾部还附着有一些尾鞘、尾丝和尾钉等。其头壳中包裹着噬菌体的基因组,而尾部的各个组件则在噬菌体感染细菌的过程中发挥作用。
痘病毒是一种具有特殊形态的体形较大的复合型病毒。其病毒基因组与结合蛋白位于称为拟核的一个中心区域。拟核圍繞在一层膜和两个未知功能的侧体中。痘病毒具有外层包膜,包膜外有一层厚的蛋白质外壳布满整个表面。痘病毒的形态有轻微的多态性,从卵状到砖块状都有。[67]
拟菌病毒(mimivirus)是目前已知最大的病毒,其衣壳直径达400nm,体积接近小型细菌,且表面布满长达100nm的蛋白质纤维丝。在电镜下观察到的拟菌病毒呈六边形,因此推测其衣壳应为二十面体对称。[68]
遗传物质
| 性质 | 参数 |
|---|---|
| 核酸类型 |
|
| 形状 |
|
| 链型 |
|
| 义 |
|
与一般的细胞生物的遗传物质为双链DNA不同的是,病毒的遗传物质(即病毒基因组)可以为DNA或RNA,可以为单链或双链。从目前已发现的病毒来看,更多的是RNA病毒;其中,植物病毒多为单链RNA病毒,而噬菌体多为双链DNA病毒。[69]不同病毒的遗传物质中的基因结构也各不相同,它们之间的差异性比动物、植物或细菌中任何一个生物域内物种间的差异性都要大。
病毒的核酸可以是环状的,如多瘤病毒,或线状的,如腺病毒。核酸的种类与其所呈现的形状无关。在RNA病毒中,病毒体中的核酸常可以分裂为多个区段,这种状态称为“分段”(segmented)。其中的每一段常常编码一个蛋白质,并且这些区段通常位于同一个衣壳中。但每一个区段并不一定要在同一个病毒体中才能使病毒整体具有感染性,雀麦花叶病毒(Brome mosaic virus)就是一个例子。[6]
病毒的核酸可以是单链或双链,也与核酸的种类无关。双链的病毒核酸是由两条互补配对的核酸链所组成,如同一个梯子。而单链的病毒核酸是一条没有配对的核酸链,如同一个梯子从中间分成两边的其中一边。一些病毒,如肝病毒科中的部分病毒,其核酸部分为单链,部分为双链。[69]
其核酸为RNA或单链DNA的病毒,其核酸链可以分为正义链或反义链,这种划分取决于其与病毒mRNA是否互补。正义病毒RNA与病毒mRNA等同,宿主细胞因此可以直接用其來翻译。反义病毒RNA与病毒mRNA互补,必须通过RNA聚合酶合成正义病毒RNA后,才能够进行翻译。单链DNA的情况与RNA相似,“编码链”(与病毒mRNA互补)为反义链(−),而“非编码链”为正义链(+)。[69]
不同病毒的核酸的大小差别很大。最小的病毒基因组分子量只有106道尔顿,编码4个蛋白质;最大的病毒基因组则有108道尔顿,编码超过100个蛋白质。[69] RNA病毒的基因组通常比DNA病毒来的小,这是由于其复制过程有更高的错误率,使得RNA病毒的大小有上限。如果超出这一上限,复制后的RNA病毒基因组会出现错误,使得导致病毒无功能或无竞争力。为了弥补这一缺陷,RNA病毒通常将自己的核酸分成多个分子,每一个分子就是一段基因组,这样也就降低了复制过程中的错误率。相比而言,由于DNA病毒具有保真度较高的复制酶,降低了复制的错误几率,因而其基因组也就更大。
遗传变异

两个不同的流感病毒株如何通过抗原转移或基因重排,形成新的病毒株。
病毒的遗传变异可以有多种机制。“遗传漂变”(genetic drift)是其中之一,即病毒DNA或RNA上单个碱基的突变。大多数这样的单点突变是无义的(或者说是沉默的),因为它们没有导致所编码的蛋白质发生变化;但有一小部分突变可能会引起进化上的优势,如产生对抗病毒药物的抵抗力。[70]抗原漂移(antigenic shift)是另一种病毒基因组的主要变化,是由遗传重组或基因重排所导致的。当流感病毒发生抗原转移后,可能会导致瘟疫。[71] RNA病毒常常以准种(quasispecies)的形式或大量同种但基因组核苷酸序列存在微小差异的病毒的形式存在。这样的准种是自然选择的主要目标。[72]
分段的基因组具有进化上的优势:同一种病毒的不同的病毒株中的分段基因组可以进行重排,重排后产生的后代病毒具有不同于上一代的独特的特征。这种基因重排又称为“病毒的有性生殖”(viral sex)。[73]
遗传重组是一条DNA链断裂后重新连接到另一条不同DNA分子末端的过程。遗传重组可以在病毒感染细胞的同时发生。对病毒进化的研究结果显示,在已研究的各种病毒中,重组发生得极为频繁。[74]而且,无论是RNA病毒还是DNA病毒,重组的发生都是非常普遍的。[75][76]
生命周期
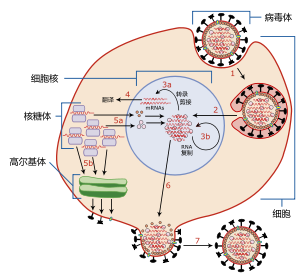
流感病毒自我复制过程简图:1.病毒体附着到宿主细胞表面并通过胞吞进入细胞;2.衣壳分解后,病毒核糖核蛋白转运入核;3a.病毒基因组转录;3b.病毒基因组复制;4.新合成的病毒mRNA出核并完成翻译;5a.合成的核蛋白入核与新复制的核酸结合;5b.合成的病毒表面蛋白进入高尔基体完成翻译后修饰并转运上膜;6.新形成的核衣壳进入细胞质并与插有病毒表面蛋白的细胞膜结合;7.新生成的病毒体通过出泡方式离开宿主细胞。
由于病毒是非细胞的,无法通过细胞分裂的方式来完成数量增长;它们是利用宿主细胞内的代谢工具来合成自身的拷贝,并完成病毒组装。不同的病毒之间生命周期的差异很大,但大致可以分为六个阶段:[77]
附着:首先是病毒衣壳蛋白与宿主细胞表面特定受体之间發生特异性结合。这种特异性决定了一种病毒的宿主范围。例如,艾滋病毒只能感染人类T细胞,因为其表面蛋白gp120能够与T细胞表面的CD4分子和受体结合。这种吸附机制通过不断的进化,使得病毒能够更特定地结合那些使它们能够完成复制过程的细胞。对于带包膜的病毒,吸附到受体上可以诱發包膜蛋白發生构象变化从而导致包膜与细胞膜發生融合。
入侵:病毒附着到宿主细胞表面之后,通过受体介导的胞吞或膜融合进入细胞,这一过程通常称为“病毒进入”(viral entry)。感染植物细胞与感染动物细胞不同,因为植物细胞有一层由纤维素形成的坚硬的细胞壁,病毒只有在细胞壁出现伤口时才能进入。[78]一些病毒,如烟草花叶病毒可以直接在植物内通过胞间连丝的孔洞从一个细胞运动到另一个细胞。[79]与植物一样,细菌也有一层细胞壁,病毒必须通过这层细胞壁才能够感染细菌。一些病毒,如噬菌体,进化出了一种感染细菌的机制,将自己的基因组注入细胞内而衣壳留在细胞外,从而减少进入细菌的阻力。[80]
脱壳:然后病毒的衣壳遭到宿主细胞或病毒自己的酶降解破坏,病毒的核酸得以释放。
合成:病毒基因组完成复制、转录(除了正义RNA病毒外)以及病毒蛋白质合成。
组装:将合成的核酸和蛋白质衣壳各部分组装在一起。在病毒颗粒完成组装之后,病毒蛋白常常会发生翻译后修饰。在诸如艾滋病毒等一些病毒中,这种修饰作用(有时称为成熟过程),可以发生在病毒从宿主细胞释放之后。[81]
释放:无包膜病毒需要在细胞裂解(通过使细胞膜发生破裂的方法)之后才能得以释放。对于包膜病毒则可以通过出泡的方式得以释放。在出泡的过程中,病毒需要从插有病毒表面蛋白的细胞膜结合,获取包膜。
DNA病毒
大多数DNA病毒基因组的复制发生在细胞核内。只要细胞表面有合适的受体,这些病毒就能够通过胞吞或膜融合的方式进入细胞。多数DNA病毒完全依赖宿主细胞的DNA和RNA的合成工具以及RNA的加工工具。而病毒基因组必须穿过核膜来获得对这些工具的控制。[82][83]
RNA病毒
RNA病毒的复制过程比较独特,由于其遗传信息保存在RNA上,因此复制过程通常发生在细胞质中。根据复制方式的不同,RNA病毒可以分为4个不同的组别(见分类一节)。RNA病毒的极性(即病毒RNA能否直接用于蛋白质合成)以及RNA是单链还是双链,很大程度上决定了它的复制机制。RNA病毒是用它们自己的RNA复制酶来对基因组进行复制。[84]
反转录病毒
反转录病毒基因组的复制是采用反转录的方式来完成的,即利用RNA模板来合成DNA。遗传物质为RNA的反转录病毒以DNA为中间物来复制其基因组,而遗传物质为DNA的反转录病毒则以RNA为中间物来复制。两类病毒都需要用到反转录酶。反转录病毒常常可以将通过反转录合成的DNA整合到宿主细胞的基因组中。能够抑制反转录酶活性的抗病毒药物(如齐多夫定和拉米夫定)可以有效地对抗反转录病毒(如艾滋病毒和包括乙肝病毒在内的肝病毒科病毒)。
命名
病毒的命名並無絕對的規則,常依病毒的型態、感染對象、最初發現地點。例如感染動植物的病毒可能依感染的對象、病徵來命名,例如麻疹病毒、狂犬病毒,以發現地點命名的包括伊波拉病毒。噬菌體的命名常依實驗室內編號命名,例如T1噬菌體。
分類
由於病毒並不像其他生物能藉由交配產生後代,因此在種別的定義上與一般生物有所不同。
ICTV分类法
國際病毒分類委員會(International Committee on Taxonomy of Viruses,簡稱ICTV)在1966年建立起了一个病毒分类的通用系统和统一的命名法则。其七届ICTV会议首次规范化了病毒物种的概念,即病毒分类的分支层次中的最低分类单元。[85]分類的主要依據是病毒顆粒的特性、抗原特性與生物特性。
到目前为止只有一小部分的病毒得到了研究,从来自人体的病毒样品中发现有20%的序列是未曾发现过的,而来自环境中(如海水、大洋沉积物等)的病毒样品则大部分的序列都是全新的。[86]
分类结构如下:
门(-viricota)
- 亚门(-viricotina)
纲(-viricetes)
目(-virales)
- 亚目(-virineae)
科(-viridae)
- 亚科(-virinae)
属(-virus)
- 亚属(-virus)
- 种
- 亚属(-virus)
- 亚科(-virinae)
- 亚目(-virineae)
- 亚门(-viricotina)
在最近(2016年)的ICTV分类中,8个目已经建立,分别是布尼亞病毒目(Bunyavirales)、有尾噬菌體目(Caudovirales)、疱疹病毒目(Herpesvirales)、线状病毒目(Ligamenvirales)、單股反鏈病毒目(Mononegavirales)、网巢病毒目(Nidovirales)、微RNA病毒目(Picornavirales)和芜菁黄花叶病毒目(Tymovirales)。分类委员会没有正式区分亚种、株系和分离株之间的区别。分类表中总共有8个目、119个科、35个亚科、725个属、4300个种以及约3000种尚未分类的病毒类型。[87][88]
巴尔的摩分类法
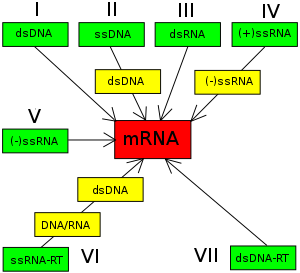
巴尔的摩分类法是基于病毒mRNA的合成方式。
诺贝尔奖获得者生物学家戴维·巴尔的摩在1970年代提出了巴尔的摩分类系统。[29][89]巴尔的摩分类法与ICTV分类法一起用于现代病毒的分类。[90][91][92]
巴尔的摩分类法是基于病毒mRNA的生成机制。在从病毒基因组到蛋白质的过程中,必须要生成mRNA来完成蛋白质合成和基因组的复制,但每一个病毒家族都采用不同的机制来完成这一过程。病毒基因组可以是单链或双链的RNA或DNA,可以有也可以没有反转录酶。而且,单链RNA病毒可以是正义(+)或反义(-)。这一分类法将病毒分为7类:
- 第一类是双链DNA病毒(如腺病毒、疱疹病毒、痘病毒)
- 第二类是单链DNA病毒(+)DNA(如小DNA病毒)
- 第三类是双链RNA病毒(如呼腸孤病毒)
- 第四类是(+)单链RNA病毒(如微小核糖核酸病毒、披盖病毒)
- 第五类是(-)单链RNA病毒(如正黏液病毒、炮彈病毒)
- 第六类是单链RNA反转录病毒(如反转录病毒)
- 第七类是双链DNA反转录病毒(如肝病毒)
举一个病毒分类的例子:水痘病毒,即带状疱疹病毒,属于疱疹病毒目,疱疹病毒科,甲型疱疹病毒亚科,水疱病毒属;同时,带状疱疹病毒是巴尔的摩分类法中的第一类,因为它是双链DNA病毒,且不含有反转录酶。
病毒與人類疾病
由病毒引起的人类疾病种类繁多。已经确定的如,感冒、流感、水痘等一般疾病,以及天花、艾滋病、SARS和禽流感等严重疾病,还有一些疾病可能是以病毒为致病因子;例如:人疱疹病毒6型与一些神经性疾病,如多发性硬化症和慢性疲勞症候群之间可能相关。此外,原本认为是马類神经系统疾病的致病因子的玻那病毒,现在发现其可能足以引起人类精神疾病。[93]病毒能够导致疾病的能力称为病毒性(virulence)。
不同的病毒有着不同的致病机制,主要取决于病毒的种类。在细胞水平上,病毒主要的破坏作用是导致细胞裂解,从而引起细胞死亡。在多细胞生物中,一旦机体内有足够多的细胞死亡,就会对机体的健康产生影响。虽然病毒可以引发疾病,却也可以无害地存在于机体内。例如,能够引起冷疮的单纯疱疹病毒可以在人体内保持休眠状态;这种状态又称为“潜伏”(latency),[94]这也是所有疱疹病毒(包括能够导致腺热的艾伯斯坦-巴尔病毒和能够导致水痘的水痘-带状疱疹病毒)的特点。进入潜伏状态的水痘-带状疱疹病毒在“苏醒”后,能够引起带状疱疹。
一些病毒能够引起慢性感染,可以在机体内不断复制而不受宿主防御系统的影响。[95]这类病毒包括乙肝病毒和丙型肝炎病毒。受到慢性感染的人群即是病毒携带者,因为他们相当于储存了保持感染性的病毒。[96]当人群中有较高比例的携带者时,这一疾病就可以发展为流行病。[97]
流行病学
病毒的流行病学是现代医学中的一个分支,主要是研究基于人的病毒的传播途径和对病毒感染的控制方法。病毒的传播可以是垂直方式,如从母亲到婴儿,或者是水平方式,即从一个人到另一个人。垂直传播的例子包括乙肝病毒和艾滋病毒,婴儿一出生就会从母亲处感染病毒;[98]另一个少见的例子是水痘-带状疱疹病毒,虽然只会引起较温和的感染反应,但对于胎儿或刚出生的婴儿却是致命的。[99]水平传播是最普遍的病毒在人群中的传播方式。病毒的传播途径包括:血液交换或性行为,如艾滋病毒、乙肝和丙肝病毒;口部的唾液交换,如人类疱疹病毒第四型;含病毒的食物或饮用水,如诺罗病毒;呼吸入以气溶胶形式存在的病毒,如流感病毒;以蚊虫为载体,通过蚊虫叮咬注入人体,如登革熱病毒。病毒感染的传播速率取决于多重因素:包括人口密度、易感染个体(例如缺乏对该病毒的免疫力)的数量[100]、医疗和天气条件等。[101]
流行病学研究是用于在病毒性疾病爆发时及时切断病毒在人群中的传染链。[102]防治措施是针对病毒的传播方式进行控制。发现病毒源和鉴定病毒是其中的重要环节。一旦鉴定了病毒,其传播就可以通过注射疫苗来阻断。当没有可用的疫苗时,改善环境卫生以及采取消毒措施也可以有效地防止病毒传播。通常受感染的病人需要与其他健康人群隔离开,而对有接触过病毒的人需要进行隔離檢疫。[103]大多数人和动物的病毒感染具有一定的潜伏期,在这一期间,不会有患病的迹象和症状。[104]病毒性疾病的潜伏期从几天到几个星期不等,但多数病毒感染的潜伏期已经得以确定。[105]在潜伏期之后,会有一段“可传染期”,即此期间受感染的个体可以将病毒传染给其他个体。[106]对于了解可传染期和潜伏期的长短对于控制疾病的爆发也非常重要。[107]当疾病爆发导致一个人群、社区或地区中有反常的高比例患病者时,这一疾病就称为流行病;如果疾病传播到世界范围则称为瘟疫。[108]
流行病和瘟疫
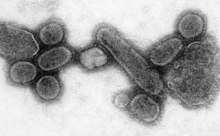
重构的西班牙流感病毒
美洲原住民的人口曾经由于欧洲殖民者带来的传染性疾病,特别是天花的蔓延而大量减少。没有详细的数据显示在哥伦布到达美洲之后有多少原住民死于外来的疾病,但估计接近总人口的70%。这一破坏性影响很大程度上帮助了欧洲殖民者对原住民进行驱赶和征服。這在當時對美洲原住民帶來了極大的恐懼。[109][110][111][112][113][114][115]
瘟疫是世界范围内的流行病。1918年的流感大爆发,即通常所说的西班牙流感,就是一种级别最高(第五级)的大规模流行病,是由一种特殊的严重致死性流感A型病毒;这一流感的感染者通常是健康的青壮年,而其他大多数流感则多发生在青少年、老人或体弱多病者中。[116]西班牙流感持续时间从1918年到1919年,过去估计死亡人数在4千万到5千万人,[117]而最新的研究结果显示这一瘟疫可能导致一亿人死亡,死亡人口占1918年全球总人口的5%。[118]
@media all and (max-width:720px){.mw-parser-output .tmulti>.thumbinner{width:100%!important;max-width:none!important}.mw-parser-output .tmulti .tsingle{float:none!important;max-width:none!important;width:100%!important;text-align:center}}


多数研究者相信艾滋病来源于二十世纪的撒哈拉以南非洲地区;[119]现在,艾滋病已经发展成为瘟疫,全球大约有3.86千万人感染了艾滋病。[120]联合国艾滋病规划署(UNAIDS)和世界卫生组织(WHO)估计自1981年6月5日发现首个病例至今,已有超过两千五百万人死于艾滋病,这使得艾滋病成为有史以来最具傷害性的瘟疫之一。[121] 2007年全年总共有2.7百万新的艾滋病感染者和2百万艾滋病相关的死亡案例。[122]
数种高致死性病毒菌株是絲狀病毒科成员。絲狀病毒是一类纤维状的病毒,可以引起病毒性出血热,其成员包括埃博拉病毒和馬爾堡病毒。馬爾堡病毒引起全世界的广泛注意是在2005年4月,安哥拉发生的一次疾病爆发。这次爆发从2004年10月一直持续到2005年。[123]伊波拉出血熱則首度出現於1976年,後造成間歇性爆發及高致死率。最近且最嚴重的一次爆發,為西非伊波拉病毒疫症。[124]
癌症
病毒是导致癌症发生的原因之一。与人类癌症相关的主要病毒有人類乳突病毒、乙肝病毒、艾伯斯坦-巴尔病毒和人类嗜T淋巴细胞病毒(human T-lymphotropic virus)。肝炎病毒可以诱发慢性病毒感染从而导致肝癌。[125][126]人类嗜T淋巴细胞病毒可以导致热带痉挛性瘫痪(tropical spastic paraparesis)和成人T细胞白血病。[127]人類乳突病毒是子宫颈癌、皮肤癌、肝门癌和阴茎癌的成因。[128]在疱疹病毒科中,卡波西肉瘤相关疱疹病毒(Kaposi's sarcoma-associated herpesvirus)能够导致卡波西氏肉瘤(Kaposi's sarcoma)和体腔淋巴瘤(body cavity lymphoma),而艾伯斯坦-巴尔病毒可以导致伯奇氏淋巴瘤(Burkitt's lymphoma)、霍奇金淋巴瘤(Hodgkin's lymphoma)、B淋巴扩增紊乱(B lymphoproliferative disorder)和鼻咽癌(nasopharyngeal carcinoma)。[129]
宿主防御机制
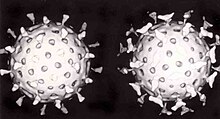
两个輪狀病毒:右侧的病毒表面结合了抗体,阻止其黏附到细胞表面,从而抑制了病毒感染。
人体抵御病毒的第一道防线是先天性免疫系统。这一免疫系统由能够抵御非特异性病毒感染的细胞和其他机制组成,即以一种通用方式来对入侵的病原体做出识别和反应,但不同于获得性免疫系统,这一免疫系统并不产生持久的或保护性的免疫。[130]
RNA干扰是对抗病毒的一种重要的先天性防御机制。[131]许多病毒需要合成双链RNA来进行复制。当这些病毒感染细胞时,它们会释放出其RNA分子,这些RNA分子立即与一种称为Dicer酶的蛋白质复合物结合,随后Dicer酶將这些RNA切为多个小的片段。一种称为RISC复合物的生化途径受到激活,开始降解病毒mRNA,使得细胞能够从病毒感染中存活下来。輪狀病毒可以逃过这层防御,一方面它进入细胞后并不完全脱壳,另一方面它可以通过病毒颗粒内衣壳上的孔洞来释放新合成的mRNA,这样其病毒基因组(为双链RNA)依然保护在病毒体内部而不会受到破坏。[132][133]
当人体的获得性免疫系统探测到病毒时,会产生特异性的抗体来与病毒结合并使其失去感染性,这种作用称为体液免疫。其中,有两类抗体非常重要。第一类称为IgM(免疫球蛋白M),它能高效地使病毒失去活性,但免疫系统的细胞产生IgM的时间只有几个星期。第二类称为IgG(免疫球蛋白G),它能够透過免疫系统不停地制造出来。IgM存在于宿主的血液中是用于急性感染的情况,而IgG的存在则表明过去曾经受到某种感染(用于防御以后的同类感染)。[134]进行免疫性测试时,通常是对体内的IgG型抗体进行测量。[135]
人体对抗病毒的第二道防线是细胞免疫(cell-mediated immunity),包括了称为T细胞的免疫细胞。人体中的细胞不断地将其内部蛋白质的片断展示在细胞表面(抗原呈递)供T细胞来进行检查,一旦T细胞识别出可能的病毒片断,那么对应的细胞就会由T杀手细胞和病毒特异性T细胞扩增所消灭。诸如巨噬细胞在内的一些细胞专门负责抗原呈递。[136]制造干扰素是一种重要的宿主防御机制。干扰素是病毒感染之后由机体所产生的一种激素,它在免疫中的作用较为复杂,可以确定的是它能够通过杀死受感染细胞及其邻近细胞来逐步阻止病毒的复制。[137]
并非所有的病毒感染都会引起保护性免疫反应。例如,艾滋病毒可以通过不断地变换其病毒体表面蛋白的氨基酸序列来逃避免疫系统的打击。这些顽固的病毒采用多种方式来逃脱免疫系统的控制,如隔离、阻断抗原呈递、产生细胞因子抗性、逃避自然杀伤细胞的作用、逃脱细胞凋亡以及抗原转移。[138]其他一些病毒,如向神经病毒(neurotropic virus),可以通过神经来传播,而在神经系统中免疫系统可能无法接触到它们。
预防与治疗
因为病毒使用了宿主细胞来进行复制并且寄居其内,因此很难用不破坏细胞的方法来杀灭病毒。现在最積極的对付病毒疾病的方法是疫苗接種来预防病毒感染或者使用抗病毒药物来降低病毒的活性以达到治疗的目的。
疫苗
疫苗接種是一种廉价而又有效的防止病毒感染的方法。早在发现病毒之前,疫苗就已经为人们用于预防病毒感染。随着疫苗接種的普及,病毒感染相关的一些疾病(如小兒麻痺、痲疹、腮腺炎和风疹)的发病率和死亡率都大幅度下降,[139]而曾经是致命疾病的天花已经绝迹。[140]目前各类疫苗可以预防超过30种对人体的病毒感染,[141]而有更多的疫苗用于防止动物受到的病毒感染。[142]疫苗的成分可以是活性降低的或死亡的病毒,也可以是病毒蛋白质(抗原)。[143]活疫苗包含了活性减弱的可致病的病毒,这样的病毒称为“减毒”病毒。虽然活性减弱,但活疫苗对于那些免疫力较弱或免疫缺陷的人可能是危险的,对他们注射活疫苗可能反而会导致疾病。[144]生物技术和基因工程用于改造病毒疫苗,改造后的疫苗(即亚单位疫苗)只含有病毒的衣壳蛋白,如乙肝疫苗。[145]由于不含有病毒核酸,因此亚单位疫苗对于免疫缺陷的病人是安全的。[146]
对于活疫苗的安全性也有一些例外,如黄热病毒疫苗,虽然是一种减毒病毒株(称为17D),却可能是目前所有疫苗中最安全和最有效的。[147]
抗病毒药物

鳥苷
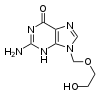
鳥苷的类似物阿昔洛韦(Aciclovir)
在过去的二十年间,抗病毒药物的发展非常迅速。艾滋病的不断蔓延推动了对抗病毒药物的需求。抗病毒药物常是核苷类似物,当病毒复制时如果将这些类似物当作核苷用于合成其基因组就会产生没有活性的病毒基因组(因为这些类似物缺少与磷相连能够相互连接形成DNA“骨架”的羟基,会造成DNA的链终止),从而抑制病毒的增殖。[148]核苷类似物作为抗病毒药物的例子包括阿昔洛韦,可用于抑制单纯疱疹病毒感染,和拉米夫定,可用于治疗艾滋病和乙型肝炎。阿昔洛韦是最早出现也是最经常指定使用的抗病毒药物。[149]
其他使用中的抗病毒药物是针对病毒生活周期的不同阶段。艾滋病毒需要依赖一种称为HIV-1蛋白酶的作用来获得完整的感染能力;而通过使用大量的蛋白酶抑制剂类的药物可以使这种酶失活。
丙型肝炎是由一种RNA病毒引起的。80%的受感染者都会形成慢性肝炎,如果不加以治疗,受感染者会一直保持感染的状态。之前有效的治疗方法是将核苷类似物利巴韦林与干扰素一起使用。现在最有效的治疗方法是直接作用于丙肝病毒蛋白酶、RNA聚合酶等病毒复制重要环节小分子药物(DAAs)[150]对于慢性乙型肝炎的病毒携带者的治疗方法也采用类似的策略。[151]
其他物种中的病毒感染
病毒可以感染所有的物种,少数病毒(如mimivirus)甚至也会受到其他特定病毒的感染[152];但特定的病毒感染物种的范围是有限的。[153]例如,植物病毒不会感染动物,而噬菌体只能感染细菌。
动物
对家畜来说,病毒是重要的致病因子;能够导致的疾病包括口蹄疫、蓝舌病等。[154]作为人类宠物的猫、狗、马等,如果没有接种疫苗,会感染一些致命病毒。例如犬小病毒(Canine parvovirus),一种小DNA病毒,其感染是导致幼犬死亡的重要原因。[155]所有的无脊椎动物都会感染病毒。例如蜜蜂会受到多种病毒的感染。[156]幸运的是,大多数病毒能够与宿主和平相处而不引起任何损害,也不导致任何疾病。[13]
植物

辣椒受到轻斑驳病毒的感染。
植物病毒的种类繁多,能够影响受感染植物的生长和繁殖。植物病毒的传播常常是由称为“载体”的生物来完成。这些载体一般为昆虫,也有部分情况下为真菌、线虫动物以及一些单细胞生物。控制针对植物的病毒感染,通常是采用消灭载体生物以及除去其他可能的病毒宿主,如杂草。[157]对于人类及其他动物来说,植物病毒是无害的,因为它们只能够在活的植物细胞内进行复制。[158]
植物具备精巧而有效的防御机制来抵抗病毒感染。其中,最为有效的机制是“抵抗基因”(R基因)。每个R基因能够抵抗一种特定病毒,主要是通过触发受感染细胞的附近细胞的死亡而产生肉眼可见的空点,从而阻止感染的扩散。[159]植物中的RNA干扰也是一种有效的防御机制。[160]当受到感染,植物常常就能够产生天然消毒剂(如水杨酸、一氧化氮和活性氧分子)来杀灭病毒。[161]
细菌
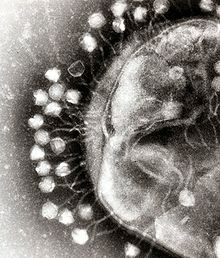
电镜照片显示了多个噬菌体附着在一个细菌的细胞壁表面。
噬菌体是病毒中最为普遍和分布最广的群体。例如,噬菌体是水体中最普遍的生物个体,在海洋中其数量可达细菌数量的十多倍,[162] 1毫升的海水中可含有约2亿5千万个噬菌体。[163]噬菌体是通过结合细菌表面的受体来感染特定的细菌。在进入细菌后的很短的时间内,有时仅仅为几分钟,细菌的聚合酶就开始将病毒mRNA翻译为蛋白质。这些病毒蛋白质有些在细菌细胞内组装成新的病毒体,有些为辅助蛋白可以帮助病毒体的组装,有些则参与细胞裂解(病毒可以产生一些酶来帮助裂解细胞膜)。噬菌体的整个感染过程非常迅速;以T4噬菌体为例,从注入病毒核酸到释放出超过300个新合成的病毒,所需的时间仅为20多分钟。[164]
细菌防御噬菌体的主要方法是合成能够降解外来DNA的酶。这些酶称为限制性内切酶,它们能够剪切噬菌体注入细菌细胞的病毒DNA。[165]细菌还含有另一个防御系统,这一系统利用CRISPR序列来保留其过去曾经遇到过的病毒的基因组片断,从而使得它们能够通过RNA干扰的方式来阻断病毒的复制。[166][167]这种遗传系统为细菌提供了一个类似于获得性免疫的机制来对抗病毒感染。
古菌
古菌也会受到一些病毒感染,主要是双链DNA病毒。这些病毒明显与其他病毒无相关性,它们具有多种特别的外形,如瓶状、钩杆状或泪滴状。[168]在嗜热古菌,特别是硫化葉菌(Sulfolobales)和熱變形菌(Thermoproteales)中的这类病毒已经获得了细致的研究。[169]古菌的病毒防御体系可能包括了RNA干扰(利用古菌基因组中所含的与病毒基因相关的重复DNA序列来进行)。[170][171]
应用
生命科学与医学
病毒对于分子生物学和细胞生物学的研究具有重要意义,因为它们提供了能够用于改造和研究细胞功能的简单系统。[172]研究和利用病毒为细胞生物学的各方面研究提供了大量有价值的信息。[173]例如,病毒在遗传学研究中的使用可用於協助人們了解分子遗传学的基本机制,包括DNA复制、转录、RNA加工、翻译、蛋白质转运以及免疫学等。
遗传学家常常用病毒作为载体将需要研究的特定基因引入细胞。这一方法对于细胞生产外源蛋白质,或是研究引入的新基因对于细胞的影响,都是非常有用的。病毒治疗法(virotherapy)也采用类似的策略,即利用病毒作为载体引入基因来治疗各种遗传性疾病,好处是可以定靶于特定的细胞和DNA。这一方法在癌症治疗和基因治療中的应用前景广阔。一些科学家已经利用噬菌体来作为抗生素的替代品,由于一些病菌的抗生素抗性的加强,人们对于这一替代方法的兴趣也不断增长。[174]
材料科学与纳米技术
目前纳米技术的发展趋势是制造多用途的病毒。从材料科学的观点来看,病毒可以視同有机纳米颗粒:它们的表面携带特定的工具用于穿过宿主细胞的壁垒。病毒的大小和形状,以及它们表面的功能基团的数量和性质,是经过精确地定义的。正因为如此,病毒在材料科学中普遍用作支架来共价连接表面修饰。病毒的一个特点是它们能够通过直接进化来改动。从生命科学发展而来的这些强大技术正在成为纳米材料制造方法的基础,远远超越了它们在生物学和医学中的应用而用于更加广泛的领域中。[175]
由于具有合适的大小、形状和明确的化学结构,病毒在纳米量级上的组织材料作為模板使用。最近的一个应用例子是利用豇豆花叶病毒颗粒来放大DNA微陣列上感应器的信号;在该应用中,病毒颗粒将用于显示信号的荧光染料分离开,从而阻止能够导致荧光淬灭的非荧光二聚体的形成。[176]另一个例子是利用豇豆花叶病毒作为纳米量级的分子电器的面板。[177]在实验室中,病毒还可以用於製造可充电电池。[178]
武器
病毒能够引起瘟疫而导致人类社会的恐慌,这种能力使得一些人企图利用病毒作为生化武器来达到常规武器所不能获得的效果。而随着臭名昭著的西班牙流感病毒在实验室中获得成功复原,对于病毒成为武器的担心不断增加。[179]另一个可能成为武器的病毒是天花病毒。天花病毒在绝迹之前曾经引起无数次的社会恐慌。目前天花病毒存在于世界上的数个安全实验室中,对于其可能成为生化武器的恐惧并非是毫无理由的。天花病毒疫苗是不安全的,在天花绝迹前,由于注射天花疫苗而患病的人数比一般患病的人数还要多,[180]而且天花疫苗目前也不再广泛生产。[181]因此,在存在如此多对于天花没有免疫力的现代人的情况下,一旦天花病毒释放出来,在病毒得到控制之前,将会有无数人患病死去。
参见
- 病毒学
- 类病毒
- 拟病毒
- 朊病毒
- 部分抗病毒药物列表
- 非细胞生物
- 人畜共通傳染病
參考文獻
引用
^ 周德庆.微生物学教程(第三版).高等教育出版社.2011.p65-66
^ Villarreal p. 3
^ 3.03.1 (英文)Norrby E. Nobel Prizes and the emerging virus concept. Arch. Virol. 2008, 153 (6): 1109–23. PMID 18446425. doi:10.1007/s00705-008-0088-8.
^ Dimmock p. 49
^ (英文)Carlson GA, Hsiao K, Oesch B, Westaway D, Prusiner SB. Genetics of prion infections 7: 61–65. 1991. PMID 1903568.
^ 6.06.16.2 Collier pp. 33–55
^ Villarreal p. 4
^ (英文)Wilson, G., "Restriction and Modification Systems," Annual Review of Genetics (1991), 25:585-627.
^ (英文)Antiviral Drugs. 2007-09-11 [2008-10-03]. (原始内容存档于2008-09-18).
^ 病毒的发现与研究历史. [2008-09-30]. (原始内容存档于2008-06-22).
^ Shors pp. 76–77
^ 12.012.112.212.3 Collier p. 3
^ 13.013.1 Dimmock p. 4
^ Dimmock p.4–5
^ Shors p. 589
^ (英文)D'Herelle F. "On an invisible microbe antagonistic toward dysenteric bacilli": brief note by Mr. F. D'Herelle, presented by Mr. Roux. 1917.. Res. Microbiol. 2007, 158 (7): 553–4. PMID 17855060.
^ (英文)Steinhardt, E; Israeli, C; and Lambert, R.A. (1913) "Studies on the cultivation of the virus of vaccinia" J. Inf Dis. 13, 294–300
^ Collier p. 4
^ (英文)Goodpasture EW, Woodruff AM, Buddingh GJ. The cultivation of vaccine and other viruses in the chorioallantoic membrane of chick embryos. Science. 1931, 74: 371–372. PMID 17810781.
^ (英文)Rosen FS (2004). "Isolation of poliovirus—John Enders and the Nobel Prize". New England Journal of Medicine, 351, pp. 1481–83 PMID 15470207
^ (英文)Nobel Lectures, Physics 1981–1990, (1993) Editor-in-Charge Tore Frängsmyr, Editor Gösta Ekspång, World Scientific Publishing Co., Singapore.
- 其实早在1887年,Buist就利用光学显微镜观察到了经过染色的牛痘苗病毒(最大的病毒之一),但当时人们还不知道牛痘苗是一种病毒。(Buist J.B. Vaccinia and Variola: a study of their life history Churchill, London)
^ (英文)Stanley WM, Loring HS (1936). "The isolation of crystalline tobacco mosaic virus protein from diseased tomato plants". Science, 83, p.85 PMID 17756690
^ (英文)Stanley WM, Lauffer MA (1939). "Disintegration of tobacco mosaic virus in urea solutions". Science 89, pp. 345–347 PMID 17788438
^ (英文)温德尔·斯坦利在诺贝尔颁奖典礼上的演讲
^ (英文)Creager AN, Morgan GJ. After the double helix: Rosalind Franklin's research on Tobacco mosaic virus. Isis. 2008年6月, 99 (2): 239–72. PMID 18702397.
^ Dimmock p. 12
^ (英文)国际病毒分类委员会(ICTV)的已发现病毒及其发现者的列表 互联网档案馆的存檔,存档日期2009-11-11.
^ Collier p. 745
^ 29.029.1 (英文)Temin HM, Baltimore D. RNA-directed DNA synthesis and RNA tumor viruses. Adv. Virus Res. 1972, 17: 129–86. PMID 4348509.
^ (英文)Barré-Sinoussi, F., Chermann, J. C., Rey, F., Nugeyre, M. T., Chamaret, S., Gruest, J., Dauguet, C., Axler-Blin, C., Vezinet-Brun, F., Rouzioux, C., Rozenbaum, W. and Montagnier, L. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science. 1983, 220 (4599): 868–871. Bibcode:1983Sci...220..868B. PMID 6189183. doi:10.1126/science.6189183.
^ (英文)2008年诺贝尔生理学与医学奖获得者
^ (英文)Iyer LM, Balaji S, Koonin EV, Aravind L. Evolutionary genomics of nucleo-cytoplasmic large DNA viruses. Virus Res. 2006年4月, 117 (1): 156–84. PMID 16494962. doi:10.1016/j.virusres.2006.01.009.
^ Villarreal p. 10
^ (英文)Liu Y, Nickle DC, Shriner D, et al (2004). "Molecular clock-like evolution of human immunodeficiency virus type 1". Virology. 10;329(1):101–8, PMID 15476878
^ Shors p. 16
^ Collier pp. 18–19
^ Shors pp. 14–16
^ Collier pp. 11–21
^ 39.039.1 Dimmock p. 16
^ Collier p. 11
^ Collier pp. 11–12
^ Shors p. 574
^ (英文)McClintock, B. The origin and behavior of mutable loci in maize. Proc Natl Acad Sci U S A. 1950年6月, 36 (6): 344–55. PMID 15430309. doi:10.1073/pnas.36.6.344.
^ Dimmock p. 55
^ Shors 551–3
^ (英文)Tsagris EM, de Alba AE, Gozmanova M, Kalantidis K. Viroids. Cell. Microbiol. 2008年9月. PMID 18764915. doi:10.1111/j.1462-5822.2008.01231.x.
^ Shors p. 492–3
^ Collier p. 777
^ Dimmock p. 55–7
^ (英文)Liberski PP. Prion diseases: a riddle wrapped in a mystery inside an enigma. Folia Neuropathol. 2008, 46 (2): 93–116. PMID 18587704.
^ Dimmock pp. 57–58
^ (英文)Lupi O, Dadalti P, Cruz E, Goodheart C. Did the first virus self-assemble from self-replicating prion proteins and RNA?. Med. Hypotheses. 2007, 69 (4): 724–30. PMID 17512677. doi:10.1016/j.mehy.2007.03.031.
^ Dimmock pp. 15–16
^ (英文)Rybicki EP (1990) "The classification of organisms at the edge of life, or problems with virus systematics." S Aft J Sci 86:182–186
^ (英文)Holmes EC. Viral evolution in the genomic age. PLoS Biol. 2007年10月, 5 (10): e278. PMC 1994994. PMID 17914905. doi:10.1371/journal.pbio.0050278.
^ (英文)Koonin EV, Senkevich TG, Dolja VV. The ancient Virus World and evolution of cells. Biol. Direct. 2006, 1: 29. PMC 1594570. PMID 16984643. doi:10.1186/1745-6150-1-29.
^ (英文)Vlassov AV, Kazakov SA, Johnston BH, Landweber LF. The RNA world on ice: a new scenario for the emergence of RNA information. J. Mol. Evol. 2005年8月, 61 (2): 264–73. PMID 16044244. doi:10.1007/s00239-004-0362-7.
^ Collier pp. 33–37
^ Collier p. 40
^ (英文)Caspar DL, Klug A. Physical principles in the construction of regular viruses. Cold Spring Harb. Symp. Quant. Biol. 1962, 27: 1–24. PMID 14019094.
^ (英文)Crick FH, Watson JD. Structure of small viruses. Nature. 1956, 177 (4506): 473–5. PMID 13309339. doi:10.1038/177473a0.
^ Collier p. 37
^ 63.063.163.263.3 (英文)Virus Structure. 2007-08-23 [2008-10-03]. (原始内容存档于2008-09-25).
^ 64.064.1 (英文)Virus Structure. 2004-10-19 [2008-10-03]. [永久失效連結]
^ Collier pp. 40, 42
^ Collier pp. 42–43
^ (英文)Long GW, Nobel J, Murphy FA, Herrmann KL, Lourie B. Experience with electron microscopy in the differential diagnosis of smallpox. Appl Microbiol. 1970年9月, 20 (3): 497–504. PMC 376966. PMID 4322005.
^ (英文)Suzan-Monti M, La Scola B, Raoult D. Genomic and evolutionary aspects of Mimivirus. Virus Research. 2006, 117 (1): 145–155. PMID 16181700. doi:10.1016/j.virusres.2005.07.011.
^ 69.069.169.269.3 Collier pp. 96–99
^ (英文)Pan XP, Li LJ, Du WB, Li MW, Cao HC, Sheng JF. Differences of YMDD mutational patterns, precore/core promoter mutations, serum HBV DNA levels in lamivudine-resistant hepatitis B genotypes B and C. J. Viral Hepat. 2007年11月, 14 (11): 767–74 [2008-09-13]. PMID 17927612. doi:10.1111/j.1365-2893.2007.00869.x. [永久失效連結]
^ (英文)Hampson AW, Mackenzie JS. The influenza viruses. Med. J. Aust. 2006年11月, 185 (10 Suppl): S39–43 [2008-09-13]. PMID 17115950.
^ (英文)Metzner KJ. Detection and significance of minority quasispecies of drug-resistant HIV-1. J HIV Ther. 2006年12月, 11 (4): 74–81. PMID 17578210.
^ (英文)Goudsmit, Jaap. Viral Sex. Oxford Univ Press, 1998.ISBN 9780195124965
^ (英文)Worobey M, Holmes EC. Evolutionary aspects of recombination in RNA viruses. J. Gen. Virol. 1999,. 80(Pt 10): 2535–43. PMID 10573145.
^ (英文)Lukashev AN. Role of recombination in evolution of enteroviruses. Rev. Med. Virol. 2005, 15 (3): 157–67. PMID 15578739. doi:10.1002/rmv.457.
^ (英文)Umene K. Mechanism and application of genetic recombination in herpesviruses. Rev. Med. Virol. 1999, 9 (3): 171–82. PMID 10479778. doi:10.1002/(SICI)1099-1654(199907/09)9:3<171::AID-RMV243>3.0.CO;2-A.
^ Collier pp. 75–91
^ Dimmock p. 70
^ (英文)Boevink P, Oparka KJ. Virus-host interactions during movement processes. Plant Physiol. 2005年8月, 138 (4): 1815–21. PMC 1183373. PMID 16172094. doi:10.1104/pp.105.066761.
^ Dimmock p. 71
^ (英文)Barman S, Ali A, Hui EK, Adhikary L, Nayak DP. Transport of viral proteins to the apical membranes and interaction of matrix protein with glycoproteins in the assembly of influenza viruses. Virus Res. 2001, 77 (1): 61–9. PMID 11451488. doi:10.1016/S0168-1702(01)00266-0.
^ Shors p. 54
^ Collier p. 78
^ Collier p. 79
^ Fields p. 27
- As defined therein, "a virus species is a polythetic class of viruses that constitute a replicating lineage and occupy a particular ecological niche". A“polythetic" class is one whose members have several properties in common, although they do not necessarily all share a single common defining one. Members of a virus species are defined collectively by a consensus group of properties. Virus species thus differ from the higher viral taxa, which are“universal”classes and as such are defined by properties that are necessary for membership.
^ (英文)Delwart EL. Viral metagenomics. Rev. Med. Virol. 2007, 17 (2): 115–31. PMID 17295196. doi:10.1002/rmv.532.
^ (英文)ICTV病毒分类表2013. [2014-01-01].
^ (英文)ICTV主物种列表2013v2. [2016-10-26]. (原始内容存档于2015-10-02).
^ (英文)Baltimore D. The strategy of RNA viruses. Harvey Lect. 1974,. 70 Series: 57–74. PMID 4377923.
^ van Regenmortel MH, Mahy BW. Emerging issues in virus taxonomy. Emerging Infect. Dis. 2004, 10 (1): 8–13. PMID 15078590.
^ (英文)Mayo MA. Developments in plant virus taxonomy since the publication of the 6th ICTV Report. International Committee on Taxonomy of Viruses. Arch. Virol. 1999, 144 (8): 1659–66. PMID 10486120. doi:10.1007/s007050050620.
^ (英文)de Villiers EM, Fauquet C, Broker TR, Bernard HU, zur Hausen H. Classification of papillomaviruses. Virology. 2004, 324 (1): 17–27. PMID 15183049. doi:10.1016/j.virol.2004.03.033.
^ (英文)Chen C, Chiu Y, Wei F, Koong F, Liu H, Shaw C, Hwu H, Hsiao K. High seroprevalence of Borna virus infection in schizophrenic patients, family members and mental health workers in Taiwan. Mol Psychiatry. 1999, 4 (1): 33–8. PMID 10089006. doi:10.1038/sj.mp.4000484.
^ (英文)Margolis TP, Elfman FL, Leib D; 等. Spontaneous reactivation of herpes simplex virus type 1 in latently infected murine sensory ganglia. J. Virol. 2007年10月, 81 (20): 11069–74 [2008-09-13]. PMC 2045564. PMID 17686862. doi:10.1128/JVI.00243-07. 引文格式1维护:显式使用等标签 (link)
^ (英文)Bertoletti A, Gehring A. Immune response and tolerance during chronic hepatitis B virus infection. Hepatol. Res. 2007,. 37 Suppl 3: S331–8. PMID 17931183. doi:10.1111/j.1872-034X.2007.00221.x.
^ (英文)Rodrigues C, Deshmukh M, Jacob T, Nukala R, Menon S, Mehta A. Significance of HBV DNA by PCR over serological markers of HBV in acute and chronic patients. Indian journal of medical microbiology. 2001, 19 (3): 141–4. PMID 17664817.
^ (英文)Nguyen VT, McLaws ML, Dore GJ. Highly endemic hepatitis B infection in rural Vietnam. Journal of Gastroenterology and Hepatology. 2007, 22: 2093. PMID 17645465. doi:10.1111/j.1440-1746.2007.05010.x.
^ (英文)Fowler MG, Lampe MA, Jamieson DJ, Kourtis AP, Rogers MF. Reducing the risk of mother-to-child human immunodeficiency virus transmission: past successes, current progress and challenges, and future directions. Am. J. Obstet. Gynecol. 2007, 197 (3 Suppl): S3–9. PMID 17825648. doi:10.1016/j.ajog.2007.06.048.
^ (英文)Sauerbrei A, Wutzler P. The congenital varicella syndrome. Journal of perinatology : official journal of the California Perinatal Association. 2000, 20 (8 Pt 1): 548–54. PMID 11190597.
^ (英文)Garnett GP. Role of herd immunity in determining the effect of vaccines against sexually transmitted disease. J. Infect. Dis. 2005年2月,. 191 Suppl 1: S97–106. PMID 15627236. doi:10.1086/425271.
^ (英文)Platonov AE. (The influence of weather conditions on the epidemiology of vector-borne diseases by the example of West Nile fever in Russia). Vestn. Akad. Med. Nauk SSSR. 2006, (2): 25–9. PMID 16544901 (俄语).
^ Shors p. 198
^ Shors pp. 199, 209
^ Shors p. 126
^ Shors pp. 193–194
^ Shors pp. 193–94
^ Shors p. 194
^ Shors pp. 192–193
^ (英文)Ranlet P. The British, the Indians, and smallpox: what actually happened at Fort Pitt in 1763?. Pa Hist. 2000, 67 (3): 427–41. PMID 17216901.
^ (英文)Van Rijn K. "Lo! The poor Indian!" colonial responses to the 1862–63 smallpox epidemic in British Columbia and Vancouver Island. Can Bull Med Hist. 2006, 23 (2): 541–60. PMID 17214129.
^ (英文)Patterson KB, Runge T. Smallpox and the Native American. Am. J. Med. Sci. 2002年4月, 323 (4): 216–22. PMID 12003378.
^ (英文)Sessa R, Palagiano C, Scifoni MG, di Pietro M, Del Piano M. The major epidemic infections: a gift from the Old World to the New?. Panminerva Med. 1999年3月, 41 (1): 78–84. PMID 10230264.
^ (英文)Bianchine PJ, Russo TA. The role of epidemic infectious diseases in the discovery of America. Allergy Proc. 1992, 13 (5): 225–32. PMID 1483570.
^ (英文)Hauptman LM. Smallpox and American Indian; Depopulation in Colonial New York. N Y State J Med. 1979年11月, 79 (12): 1945–9. PMID 390434.
^ (英文)Fortuine R. Smallpox decimates the Tlingit(1787). Alaska Med. 1988, 30 (3): 109. PMID 3041871.
^ Collier pp. 409–415
^ (英文)Patterson KD, Pyle GF. The geography and mortality of the 1918 influenza pandemic. Bull Hist Med. 1991.Spring, 65 (1): 4–21. PMID 2021692. 请检查|date=中的日期值 (帮助)
^ (英文)Johnson NP, Mueller J. Updating the accounts: global mortality of the 1918–1920 "Spanish" influenza pandemic. Bull Hist Med. 2002, 76 (1): 105–15 [2008-09-13]. PMID 11875246. doi:10.1353/bhm.2002.0022.
^ (英文)Gao, Feng; Bailes, Elizabeth; Robertson, David L.; Chen, Yalu; Rodenburg, Cynthia M.; Michael, Scott F.; Cummins, Larry B.; Arthur, Larry O.; Peeters, Martine; Shaw, George M.; Sharp, Paul M.; Hahn, Beatrice H. Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature. 1999, 397 (6718): 436–441. ISSN 0028-0836. doi:10.1038/17130.
^ Shors p. 447
^ (英文)Mawar N, Saha S, Pandit A, Mahajan U. The third phase of HIV pandemic: social consequences of HIV/AIDS stigma & discrimination & future needs (PDF). Indian J. Med. Res. 2005年12月, 122 (6): 471–84 [2008-09-13]. PMID 16517997. (原始内容 (PDF)存档于2016-03-04).
^ (英文)Status of the global HIV epidemic (PDF). UNAIDS. 2008 [2008-09-15]. (原始内容 (PDF)存档于2015-11-22).
^ (英文)Towner JS, Khristova ML, Sealy TK; 等. Marburgvirus genomics and association with a large hemorrhagic fever outbreak in Angola. J. Virol. 2006年7月, 80 (13): 6497–516. PMC 1488971. PMID 16775337. doi:10.1128/JVI.00069-06. 引文格式1维护:显式使用等标签 (link)
^ Statement on the Meeting of the International Health Regulations Emergency Committee Regarding the 2014 Ebola Outbreak in West Africa. World Health Organization. 2014-08-08 [2014-08-08].
^ (英文)Koike K. Hepatitis C virus contributes to hepatocarcinogenesis by modulating metabolic and intracellular signalling pathways. J. Gastroenterol. Hepatol. 2007,. 22 Suppl 1: S108–11. PMID 17567457. doi:10.1111/j.1440-1746.2006.04669.x.
^ (英文)Hu J, Ludgate L. HIV-HBV and HIV-HCV coinfection and liver cancer development. Cancer Treat. Res. 2007, 133: 241–52. PMID 17672044. doi:10.1007/978-0-387-46816-7_9.
^ (英文)Bellon M, Nicot C. Telomerase: a crucial player in HTLV-I-induced human T-cell leukemia. Cancer genomics & proteomics. 2007, 4 (1): 21–5. PMID 17726237.
^ (英文)Schiffman M, Castle PE, Jeronimo J, Rodriguez AC, Wacholder S. Human papillomavirus and cervical cancer. Lancet. 2007, 370 (9590): 890–907. PMID 17826171. doi:10.1016/S0140-6736(07)61416-0.
^ (英文)Klein E, Kis LL, Klein G. Epstein-Barr virus infection in humans: from harmless to life endangering virus-lymphocyte interactions. Oncogene. 2007, 26 (9): 1297–305. PMID 17322915. doi:10.1038/sj.onc.1210240.
^ (英文)Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walters, Peter. Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. 2002. ISBN 978-0-8153-3218-3.
^ (英文)Ding SW, Voinnet O. Antiviral immunity directed by small RNAs. Cell. 2007, 130 (3): 413–26 [2008-09-13]. PMID 17693253. doi:10.1016/j.cell.2007.07.039.
^ (英文)Patton JT, Vasquez-Del Carpio R, Spencer E. Replication and transcription of the rotavirus genome. Curr. Pharm. Des. 2004, 10 (30): 3769–77. PMID 15579070. doi:10.2174/1381612043382620.
^ (英文)Jayaram H, Estes MK, Prasad BV. Emerging themes in rotavirus cell entry, genome organization, transcription and replication. Virus Res. 2004, 101 (1): 67–81. PMID 15010218. doi:10.1016/j.virusres.2003.12.007.
^ (英文)Greer S, Alexander GJ. Viral serology and detection. Baillieres Clin. Gastroenterol. 1995年12月, 9 (4): 689–721. PMID 8903801. doi:10.1016/0950-3528(95)90057-8.
^ (英文)Matter L, Kogelschatz K, Germann D. Serum levels of rubella virus antibodies indicating immunity: response to vaccination of subjects with low or undetectable antibody concentrations. J. Infect. Dis. 1997年4月, 175 (4): 749–55. PMID 9086126. doi:10.1086/513967.
^ (英文)Cascalho M, Platt JL. Novel functions of B cells. Crit. Rev. Immunol. 2007, 27 (2): 141–51. PMID 17725500.
^ (英文)Le Page C, Génin P, Baines MG, Hiscott J. Interferon activation and innate immunity. Rev Immunogenet. 2000, 2 (3): 374–86. PMID 11256746.
^ (英文)Hilleman MR. Strategies and mechanisms for host and pathogen survival in acute and persistent viral infections. Proc. Natl. Acad. Sci. U.S.A. 2004年10月,. 101 Suppl 2: 14560–6. PMC 521982. PMID 15297608. doi:10.1073/pnas.0404758101.
^ (英文)Asaria P, MacMahon E. Measles in the United Kingdom: can we eradicate it by 2010?. BMJ. 2006, 333 (7574): 890–5. PMID 17068034. doi:10.1136/bmj.38989.445845.7C.
^ (英文)Lane JM. Mass vaccination and surveillance/containment in the eradication of smallpox. Curr. Top. Microbiol. Immunol. 2006, 304: 17–29. PMID 16989262. doi:10.1007/3-540-36583-4_2.
^ (英文)Arvin AM, Greenberg HB. New viral vaccines. Virology. 2006, 344 (1): 240–9. PMID 16364754. doi:10.1016/j.virol.2005.09.057.
^ (英文)Pastoret PP, Schudel AA, Lombard M. Conclusions--future trends in veterinary vaccinology. Rev. - Off. Int. Epizoot. 2007, 26 (2): 489–94, 495–501, 503–9. PMID 17892169.
^ (英文)Palese P. Making better influenza virus vaccines?. Emerging Infect. Dis. 2006, 12 (1): 61–5. PMID 16494719.
^ (英文)Thomssen R. Live attenuated versus killed virus vaccines. Monographs in allergy. 1975, 9: 155–76. PMID 1090805.
^ (英文)McLean AA. Development of vaccines against hepatitis A and hepatitis B. Rev. Infect. Dis. 1986, 8 (4): 591–8. PMID 3018891.
^ (英文)Casswall TH, Fischler B. Vaccination of the immunocompromised child. Expert review of vaccines. 2005, 4 (5): 725–38. PMID 16221073. doi:10.1586/14760584.4.5.725.
^ (英文)Barnett ED, Wilder-Smith A, Wilson ME. Yellow fever vaccines and international travelers. Expert Rev Vaccines. 2008年7月, 7 (5): 579–87. PMID 18564013. doi:10.1586/14760584.7.5.579.
^ (英文)Magden J, Kääriäinen L, Ahola T. Inhibitors of virus replication: recent developments and prospects. Appl. Microbiol. Biotechnol. 2005, 66 (6): 612–21. PMID 15592828. doi:10.1007/s00253-004-1783-3.
^ (英文)Mindel A, Sutherland S. Genital herpes - the disease and its treatment including intravenous acyclovir. J. Antimicrob. Chemother. 1983,. 12 Suppl B: 51–9. PMID 6355051.
^ (英文)Witthöft T, Möller B, Wiedmann KH; 等. Safety, tolerability and efficacy of peginterferon alpha-2a and ribavirin in chronic hepatitis C in clinical practice: The German Open Safety Trial. J. Viral Hepat. 2007, 14 (11): 788–96. PMC 2156112. PMID 17927615. doi:10.1111/j.1365-2893.2007.00871.x. 引文格式1维护:显式使用等标签 (link)
^ (英文)Rudin D, Shah SM, Kiss A, Wetz RV, Sottile VM. Interferon and lamivudine vs. interferon for hepatitis B e antigen-positive hepatitis B treatment: meta-analysis of randomized controlled trials. Liver Int. 2007, 27 (9): 1185–93. PMC 2156150. PMID 17919229. doi:10.1111/j.1478-3231.2007.01580.x.
^ (英文)La Scola B, Desnues C, Pagnier I, Robert C, Barrassi L, Fournous G, Merchat M, Suzan-Monti M, Forterre P, Koonin E, Raoult D. The virophage as a unique parasite of the giant mimivirus. Nature. 2008, 455 (7209): 100–4. PMID 18690211.
^ Dimmock p. 3
^ (英文)Goris N, Vandenbussche F, De Clercq K. Potential of antiviral therapy and prophylaxis for controlling RNA viral infections of livestock. Antiviral Res. 2008年4月, 78 (1): 170–8. PMID 18035428. doi:10.1016/j.antiviral.2007.10.003.
^ (英文)Carmichael L. An annotated historical account of canine parvovirus. J. Vet. Med. B Infect. Dis. Vet. Public Health. 2005, 52 (7–8): 303–11. PMID 16316389.
^ (英文)Chen YP, Zhao Y, Hammond J, Hsu H, Evans JD, Feldlaufer MF. Multiple virus infections in the honey bee and genome divergence of honey bee viruses. Journal of Invertebrate Pathology. October–November 2004, 87 (2–3): 84–93. PMID 15579317.
^ Shors p. 584
^ Shors pp. 562–587
^ (英文)Dinesh-Kumar SP, Wai-Hong Tham, Baker BJ (2000). "Structure—function analysis of the tobacco mosaic virus resistance gene N". PNAS 97, 14789-94 PMID 11121079
^ Shors pp. 573–576
^ (英文)Soosaar JL, Burch-Smith TM, Dinesh-Kumar SP (2005). "Mechanisms of plant resistance to viruses". Nat. Rev. Microbiol. 3, pp. 789–98 PMID 16132037
^ (英文)Wommack KE, Colwell RR. Virioplankton: viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000年3月, 64 (1): 69–114. PMC 98987. PMID 10704475.
^ (英文)Bergh O, Børsheim KY, Bratbak G, Heldal M. High abundance of viruses found in aquatic environments. Nature. 1989年8月, 340 (6233): 467–8. PMID 2755508. doi:10.1038/340467a0.
^ Shors pp. 595–97
^ (英文)Bickle TA, Krüger DH. Biology of DNA restriction. Microbiol. Rev. 1993年6月, 57 (2): 434–50. PMC 372918. PMID 8336674.
^ (英文)Barrangou R, Fremaux C, Deveau H; 等. CRISPR provides acquired resistance against viruses in prokaryotes. Science (journal). 2007年3月, 315 (5819): 1709–12. PMID 17379808. doi:10.1126/science.1138140. 引文格式1维护:显式使用等标签 (link)
^ (英文)Brouns SJ, Jore MM, Lundgren M; 等. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science (journal). 2008年8月, 321 (5891): 960–4. PMID 18703739. doi:10.1126/science.1159689. 引文格式1维护:显式使用等标签 (link)
^ (英文)Prangishvili D, Forterre P, Garrett RA. Viruses of the Archaea: a unifying view. Nat. Rev. Microbiol. 2006, 4 (11): 837–48. PMID 17041631. doi:10.1038/nrmicro1527.
^ (英文)Prangishvili D, Garrett RA. Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses. Biochem. Soc. Trans. 2004, 32 (Pt 2): 204–8. PMID 15046572. doi:10.1042/BST0320204.
^ (英文)Mojica FJ, Díez-Villaseñor C, García-Martínez J, Soria E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 2005, 60 (2): 174–82. PMID 15791728. doi:10.1007/s00239-004-0046-3.
^ (英文)Makarova KS, Grishin NV, Shabalina SA, Wolf YI, Koonin EV. A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action. Biol. Direct. 2006, 1: 7. PMID 16545108. doi:10.1186/1745-6150-1-7.
^ Collier p.8
^ (英文)Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James.Viruses:Structure, Function, and Uses Retrieved on September 16 2008
^ (英文)Matsuzaki S, Rashel M, Uchiyama J; 等. Bacteriophage therapy: a revitalized therapy against bacterial infectious diseases. J. Infect. Chemother. 2005, 11 (5): 211–9. PMID 16258815. doi:10.1007/s10156-005-0408-9. 引文格式1维护:显式使用等标签 (link)
^ (英文)Fischlechner M, Donath E. Viruses as Building Blocks for Materials and Devices. Angewandte Chemie International Edition. 2007, 46: 3184. doi:10.1002/anie.200603445.
^ (英文)Soto CM, Blum AS, Vora GJ; 等. Fluorescent signal amplification of carbocyanine dyes using engineered viral nanoparticles. J. Am. Chem. Soc. 2006年4月, 128 (15): 5184–9. PMID 16608355. doi:10.1021/ja058574x. 引文格式1维护:显式使用等标签 (link)
^ (英文)Blum AS, Soto CM, Wilson CD, et al (2005). "An Engineered Virus as a Scaffold for Three-Dimensional Self-Assembly on the Nanoscale". Small, 7, 702.
^ (英文)Lauren Cahoon. Bugs Build Batteries. ScienceNOW Daily News. 2009-04-03. 使用|accessdate=需要含有|url=(帮助)
^ Shors p. 331
^ (英文)Aragón TJ, Ulrich S, Fernyak S, Rutherford GW. Risks of serious complications and death from smallpox vaccination: a systematic review of the United States experience, 1963–1968. BMC public health. 2003, 3: 26. PMID 12911836. doi:10.1186/1471-2458-3-26.
^ (英文)Weiss MM, Weiss PD, Mathisen G, Guze P. Rethinking smallpox. Clin. Infect. Dis. 2004, 39 (11): 1668–73. PMID 15578369. doi:10.1086/425745.
书籍
.mw-parser-output .refbegin{font-size:90%;margin-bottom:0.5em}.mw-parser-output .refbegin-hanging-indents>ul{list-style-type:none;margin-left:0}.mw-parser-output .refbegin-hanging-indents>ul>li,.mw-parser-output .refbegin-hanging-indents>dl>dd{margin-left:0;padding-left:3.2em;text-indent:-3.2em;list-style:none}.mw-parser-output .refbegin-100{font-size:100%}
(英文)Collier, Leslie; Balows, Albert; Sussman, Max (1998) Topley and Wilson's Microbiology and Microbial Infections ninth edition, Volume 1, Virology, volume editors: Mahy, Brian and Collier, Leslie. Arnold. ISBN 978-0-340-66316-5
(英文)Dimmock, N.J; Easton, Andrew J; Leppard, Keith (2007) Introduction to Modern Virology sixth edition, Blackwell Publishing, ISBN 978-1-4051-3645-7
(英文)Knipe, David M; Howley, Peter M; Griffin, Diane E; Lamb, Robert A; Martin, Malcolm A; Roizman, Bernard; Straus Stephen E. (2007) Fields Virology Lippincott Williams & Wilkins. ISBN 978-0-7817-6060-7
(英文)Villarreal, Luis P. (2005) Viruses and the Evolution of Life ASM Press, ISBN 978-1-55581-309-3
(英文)Shors, Teri (2008). Understanding Viruses. Jones and Bartlett Publishers. ISBN 978-0-7637-2932-5
延伸阅读
莽克强. 《基础病毒学》. 化学工业出版社. 2005年. ISBN 7502569138.
张忠信. 《病毒分类学》. 高等教育出版社. 2006年. ISBN 7040192519.
Sompayrac, L. 《病毒学概览》. 化学工业出版社. 2005年. ISBN 7-5025-7698-3.
外部链接
维基共享资源中相关的多媒体资源:病毒 |
维基物种中的分类信息:病毒 |
(英文)病毒简介
(英文)病毒名称列表
|

