線粒體
body.skin-minerva .mw-parser-output table.infobox caption{text-align:center}
| 「線粒體」的各地常用別名 | |
|---|---|
 透射电子显微镜(TEM)下哺乳动物肺组织中的两个线粒体。 | |
| 中国大陸 | 线粒体 |
| 臺灣 | 粒線體 |
| 港澳 | 線粒體 |

线粒体简要图例

線粒體的示意圖:(1)內膜,(2)外膜,(3)嵴,(4)基質
| 细胞生物学 | |
|---|---|
动物细胞 | |
 动物细胞的基本结构:
|
线粒体(mitochondrion)是一种存在于大多数真核细胞中的由两层膜包被的细胞器,[1]直径在0.5到10微米左右。除了溶组织内阿米巴、篮氏贾第鞭毛虫以及几种微孢子虫外,大多数真核细胞或多或少都拥有线粒体,但它们各自拥有的线粒体在大小、数量及外观等方面上都有所不同。[2]这种细胞器拥有自身的遗传物质和遗传体系,但因其基因组大小有限,所以线粒体是一种半自主细胞器。线粒体是细胞内氧化磷酸化和合成三磷酸腺苷(ATP)的主要场所,为细胞的活动提供了化学能量,所以有“細胞的發電站”(the powerhouse of the cell)之称。[3]除了为细胞供能外,线粒体还参与诸如细胞分化、细胞信息传递和细胞凋亡等过程,并拥有调控细胞生长和细胞周期的能力。[4]
英文中的“线粒体”(mitochondrion,复数形式为“mitochondria”)一词是由希腊语中的“线”(“μίτος”或“mitos”)和“颗粒”(“χονδρίον”或“chondrion”)组合而成的。在“线粒体”这一名称出现前后,“粒体”“球状体”等众多名字曾先后或同时被使用。[5][注 1]
目录
1 概况
2 发现及研究的历史
3 结构
3.1 外膜
3.2 膜间隙
3.3 内膜
3.4 嵴
3.5 基质
4 功能
4.1 能量转化
4.1.1 三羧酸循環
4.1.2 氧化磷酸化
4.2 储存钙离子
4.3 其他功能
5 起源假说
5.1 内共生假说
5.2 非内共生假说
6 遗传学
6.1 基因组
6.2 遗传密码
6.3 分裂与融合
6.4 群体遺傳學
7 机能障碍与疾病
8 粒線體缺陷疾病
9 参见
10 注释
11 参考文献
12 外部連結
概况
- 大小
线粒体的直径一般为0.5-1.0μm,长1.5-3.0μm,在光学显微镜下可见。[6]在动物细胞中,线粒体大小受细胞代谢水平限制。[7]不同组织在不同条件下可能产生体积异常膨大的线粒体,称为“巨线粒体”(megamitochondria):胰脏外分泌细胞中可长达10-20μm;神经元胞体中的线粒体尺寸差异很大,有的也可能长达10μm;人类成纤维细胞的线粒体则更长,可达40μm。[8]有研究表明在低氧气分压的环境中,某些如烟草的植物的线粒体能可逆地变为巨线粒体,长度可达80μm,并形成网络。[9]
- 形状
线粒体一般呈短棒状或圆球状,但因生物种类和生理状态而异,还可呈环状、线状、哑铃状、分杈状、扁盘状或其它形状。成型蛋白(shape-forming protein)介导线粒体以不同方式与周围的细胞骨架接触或在线粒体的两层膜间形成不同的连接可能是线粒体在不同细胞中呈现出不同形态的原因。[10]
- 数量
不同生物的不同组织中线粒体数量的差异是巨大的。[11]有许多细胞拥有多达数千个的线粒体(如肝脏细胞中有1000-2000个线粒体),而一些细胞则只有一个线粒体(如酵母菌细胞的大型分支线粒体)。大多数哺乳动物的成熟红细胞不具有线粒体。[12]一般来说,细胞中线粒体数量取决于该细胞的代谢水平,代谢活动越旺盛的细胞线粒体越多。
- 分布
线粒体分布方向与微管一致,[13]通常分布在细胞功能旺盛的区域:如在肾脏细胞中靠近微血管,呈平行或栅状排列;在肠表皮细胞中呈两极分布,集中在顶端和基部;在精子中分布在鞭毛中区。在卵母细胞体外培养中,随着细胞逐渐成熟,线粒体会由在细胞周边分布发展成均匀分布。[14][15]线粒体在细胞质中能以微管为导轨、由马达蛋白提供动力向功能旺盛的区域迁移。
- 组成
线粒体的化学组分主要包括水、蛋白质和脂质,此外还含有少量的辅酶等小分子及核酸。蛋白质占线粒体干重的65-70%。线粒体中的蛋白质既有可溶的也有不溶的。可溶的蛋白质主要是位于线粒体基质的酶和膜的外周蛋白;不溶的蛋白质构成膜的本体,其中一部分是镶嵌蛋白,也有一些是酶。线粒体中脂类主要分布在两层膜中,占干重的20-30%。在线粒体中的磷脂占总脂质的3/4以上。同种生物不同组织线粒体膜中磷脂的量相对稳定。[16]含丰富的心磷脂和较少的胆固醇是线粒体在组成上与细胞其他膜结构的明显差别。
- 结构
线粒体由外至内可划分为线粒体外膜(OMM)、线粒体膜间隙、线粒体内膜(IMM)和线粒体基质四个功能区。处于线粒体外侧的膜彼此平行,都是典型的单位膜。其中,线粒体外膜较光滑,起细胞器界膜的作用;线粒体内膜则向内皱褶形成线粒体嵴,负担更多的生化反应。这两层膜将线粒体分出两个区室,位于两层线粒体膜之间的是线粒体膜间隙,被线粒体内膜包裹的是线粒体基质。
发现及研究的历史

阿尔伯特·冯·科立克,瑞士科学家,线粒体的发现者。
线粒体的研究是从19世纪50年代末开始的。
1857年,瑞士解剖学家及生理学家阿尔伯特·冯·科立克在肌肉细胞中发现了颗粒状结构。[17]另外的一些科学家在其他细胞中也发现了同样的结构,证实了科立克的发现。德国病理学家及组织学家理查德·阿尔特曼将这些颗粒命名为“原生粒”(bioblast)并于1886年发明了一种鉴别这些颗粒的染色法。阿尔特曼猜测这些颗粒可能是共生于细胞内的独立生活的细菌。[18]
1898年,德国科学家卡尔·本达因这些结构时而呈线状时而呈颗粒状[注 2],所以用希腊语中“线”和“颗粒”对应的两个词——“mitos”和“chondros”——组成“mitochondrion”来为这种结构命名,这个名称被沿用至今。[19]一年后,美国化学家莱昂诺尔·米歇利斯开发出用具有还原性的健那绿染液为线粒体染色的方法,并推断线粒体参与某些氧化反应。[20]这一方法于1900年公布,并由美国细胞学家埃德蒙·文森特·考德里推广。[21]德国生物化学家奥托·海因里希·沃伯格成功完成线粒体的粗提取且分离得到一些催化与氧有关的反应的呼吸酶,并提出这些酶能被氰化物(如氢氰酸)抑制的猜想。[22]
英国生物学家大卫·基林在1923年至1933年这十年间对线粒体内的氧化还原链(redox chain)的物质基础进行探索,辨别出反应中的电子载体——细胞色素。[23]
沃伯格于1931年因“发现呼吸酶的性质及作用方式”被授予诺贝尔生理学或医学奖。[24]
结构

线粒体剖面图
外膜
线粒体外膜是位于线粒体最外围的一层单位膜,厚度约为6-7nm。其中磷脂与蛋白质的质量为0.9:1,与真核细胞细胞膜的同一比例相近。线粒体外膜中酶的含量相对较少,其标志酶为单胺氧化酶。线粒体外膜包含称为“孔蛋白”的整合蛋白,其内部通道宽約2-3nm,这使线粒体外膜对分子量小于5000Da的分子完全通透。分子量大于上述限制的分子则需拥有一段特定的信号序列以供识别并通过外膜转运酶(translocase of the outer membrane,TOM)的主动运输来进出线粒体。[25]
线粒体外膜主要参与诸如脂肪酸链延伸、肾上腺素氧化以及色氨酸生物降解等生化反应,它也能同时对那些将在线粒体基质中进行彻底氧化的物质先行初步分解。细胞凋亡过程中,线粒体外膜对多种存在于线粒体膜间隙中的蛋白的通透性增加,[26]使致死性蛋白进入细胞质基质,促进了细胞凋亡。[27]高分辨三维X射线摄影可见内质网及线粒体之间的有20%膜是紧密接触的,[28]在这些接触位点上线粒体外膜与内质网膜通过某些蛋白质相连,形成称为“线粒体结合内质网膜”(mitochondria-associated ER-membrane,MAM)的结构。该结构在脂质的相互交换和线粒体与内质网间的钙离子信号传导等过程中都有重要作用。[29]
膜间隙
线粒体膜间隙是线粒体外膜与线粒体内膜之间的空隙,宽约6-8nm,其中充满无定形液体。由于线粒体外膜含有孔蛋白,通透性较高,而线粒体内膜通透性较低,所以线粒体膜间隙内容物的组成与细胞质基质十分接近,含有众多生化反应底物、可溶性的酶和辅助因子等。线粒体膜间隙中还含有比细胞质基质中浓度更高的腺苷酸激酶、单磷酸激酶和二磷酸激酶等激酶,其中腺苷酸激酶是线粒体膜间隙的标志酶。线粒体膜间隙中存在的蛋白质可统称为“线粒体膜间隙蛋白质”,这些蛋白质全部在细胞质基质中合成。[30]
内膜
线粒体内膜是位于线粒体外膜内侧、包裹着线粒体基质的单位膜。线粒体内膜中蛋白质与磷脂的质量比约为0.7:0.3,并含有大量的心磷脂(心磷脂常为细菌细胞膜的成分[31])。线粒体内膜的某些部分会向线粒体基质折叠形成线粒体嵴。线粒体内膜的标志酶是细胞色素氧化酶。
线粒体内膜含有電子傳遞鏈(ETC)以及比外膜更多的蛋白质(超过151种,约占线粒体所含所有蛋白质的五分之一),所以承担着更复杂的生化反应。存在于线粒体内膜中的几类蛋白质主要负责以下生理过程:特异性载体运输磷酸、谷氨酸、鸟氨酸、各种离子及核苷酸等代谢产物和中间产物;内膜转运酶(translocase of the inner membrane,TIM)运输蛋白质;参与氧化磷酸化中的氧化还原反应;参与ATP的合成;控制线粒体的分裂与融合。[32]
嵴
线粒体嵴简称“嵴”,是线粒体内膜向线粒体基质折褶形成的一种结构。线粒体嵴的形成增大线粒体内膜的表面积。在不同种类的细胞中,线粒体嵴的数目、形态和排列方式可能有较大差别。线粒体嵴主要有几种排列方式,分别称为“片状嵴”(lamellar cristae)、“管状嵴”(tubular cristae)和“泡状嵴”(vesicular cristae)。[33]片状排列的线粒体嵴主要出现在高等动物细胞的线粒体中,这些片状嵴多数垂直于线粒体长轴;管状排列的线粒体嵴则主要出现在原生动物和植物细胞的线粒体中。有研究发现,睾丸间质细胞中既存在层状嵴也存在管状嵴。[34]线粒体嵴上有许多有柄小球体,即线粒体基粒,基粒中含有ATP合酶,能利用呼吸链产生的能量合成三磷酸腺苷。所以需要较多能量的细胞,线粒体嵴的数目一般也较多。但某些形态特殊的线粒体嵴由于没有ATP合酶,所以不能合成ATP。[35]
基质
线粒体基质是线粒体中由线粒体内膜包裹的内部空间,其中含有参与三羧酸循环、脂肪酸氧化、氨基酸降解等生化反应的酶等众多蛋白质,所以较细胞质基质黏稠。[注 3][36]苹果酸脱氢酶是线粒体基质的标志酶。线粒体基质中一般还含有线粒体自身的DNA(即线粒体DNA)、RNA和核糖体(即线粒体核糖体)。
线粒体DNA是线粒体中的遗传物质,呈双链环状,并可与多种蛋白质结合成高度紧密的线粒体拟核。一个线粒体中可有一个或数个线粒体DNA分子。线粒体RNA是线粒体DNA的表达产物,RNA编辑也普遍存在于线粒体RNA中,是线粒体产生功能蛋白所必不可少的过程。[37]线粒体核糖体是存在于线粒体基质内的一种核糖体,负责完成线粒体内进行的翻译工作。线粒体核糖体的沉降系数介干55S-56S之间。一般的线粒体核糖体由28S核糖体亚基(小亚基)和39S核糖体亚基(大亚基)组成。[38]在这类核糖体中,rRNA约占25%,核糖体蛋白质约占75%。线粒体核糖体是已发现的蛋白质含量最高的一类核糖体。线粒体基质中存在的蛋白质统称为“线粒体基质蛋白质”,包括DNA聚合酶、RNA聚合酶、柠檬酸合成酶以及三羧酸循环酶系中的酶类。大部分线粒体基质蛋白是由核基因编码的。线粒体基质蛋白不一定只在线粒体基质中表达,它们也可以在线粒体外表达。[39]
功能
能量转化
线粒体是真核生物进行氧化代谢的部位,是糖类、脂肪和氨基酸最终氧化释放能量的场所。线粒体负责的最终氧化的共同途径是三羧酸循环与氧化磷酸化,分别对应有氧呼吸的第二、三阶段。[注 4]细胞质基质中完成的糖酵解和在线粒体基质中完成的三羧酸循环在会产还原型烟酰胺腺嘌呤二核苷酸(reduced nicotinarnide adenine dinucleotide,NADH)和还原型黄素腺嘌呤二核苷酸(reduced flavin adenosine dinucleotide,FADH2)等高能分子,而氧化磷酸化这一步骤的作用则是利用这些物质还原氧气释放能量合成ATP。在有氧呼吸过程中,1分子葡萄糖经过糖酵解、三羧酸循环和氧化磷酸化将能量释放后,可产生30-32分子ATP(考虑到将NADH运入线粒体可能需消耗2分子ATP)。[40]如果细胞所在环境缺氧,则会转而进行无氧呼吸。此时,糖酵解产生的丙酮酸便不再进入线粒体内的三羧酸循环,而是继续在细胞质基质中反应(被NADH还原成乙醇或乳酸等发酵产物),但不产生ATP。所以在无氧呼吸过程中,1分子葡萄糖只能在第一阶段产生2分子ATP。
三羧酸循環
糖酵解中生成的每分子丙酮酸会被主动运输转运穿过线粒体膜。进入线粒体基质后,丙酮酸会被氧化,并与辅酶A结合生成CO2、还原型辅酶Ⅰ和乙酰辅酶A。乙酰辅酶A是三羧酸循环(也称为「柠檬酸循环」或「Krebs循环」)的初级底物。参与该循环的酶除位于线粒体内膜的琥珀酸脱氢酶外都游离于线粒体基质中。[41]在三羧酸循环中,每分子乙酰辅酶A被氧化的同时会产生起始电子传递链的还原型辅因子(包括3分子NADH和1分子FADH2)以及1分子三磷酸鸟苷(GTP)。
氧化磷酸化
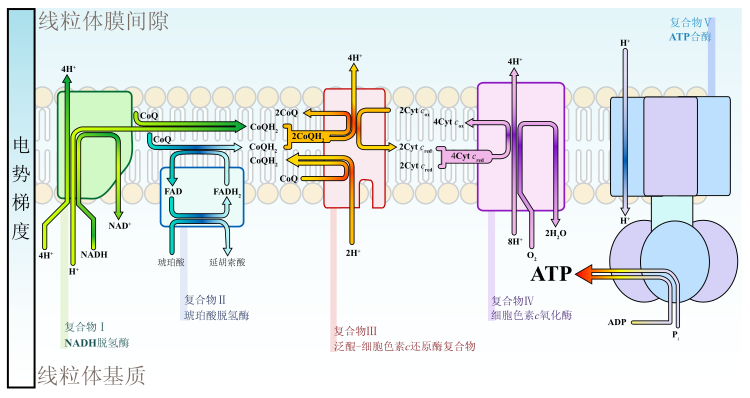
NADH和FADH2等具有还原性的分子(在细胞质基质中的还原当量可从由逆向转运蛋白构成的苹果酸-天冬氨酸穿梭系统或通过磷酸甘油穿梭作用进入电子传递链)在电子传递链里面经过几步反应最终将氧气还原并释放能量,其中一部分能量用于生成ATP,其余则作为热能散失。在线粒体内膜上的酶复合物(NADH-泛醌还原酶、泛醌-细胞色素c还原酶、细胞色素c氧化酶)利用过程中释放的能量将质子逆浓度梯度泵入线粒体膜间隙。虽然这一过程是高效的,但仍有少量电子会过早地还原氧气,形成超氧化物等活性氧(ROS),这些物质能引起氧化应激反应使线粒体性能发生衰退。[42]
当质子被泵入线粒体膜间隙后,线粒体内膜两侧便建立起了电化学梯度,质子就会有顺浓度梯度扩散的趋势。质子唯一的扩散通道是ATP合酶(呼吸链复合物V)。当质子通过复合物从膜间隙回到线粒体基质时,电势能被ATP合酶用于将ADP和磷酸合成ATP。这个过程被称为“化学渗透”,是一种协助扩散。彼得·米切尔就因为提出了这一假说而获得了1978年诺贝尔奖。1997年诺贝尔奖获得者保罗·博耶和約翰·沃克阐明了ATP合酶的机制。
储存钙离子

软骨细胞中线粒体(字母M附近的黑斑)经钙离子染料染色后得到的电镜照片。
线粒体可以储存钙离子,可以和内质网、细胞外基质等结构协同作用,[43]从而控制细胞中的钙离子浓度的动态平衡。[44]线粒体迅速吸收钙离子的能力使其成为细胞中钙离子的缓冲区。[45]在线粒体内膜膜电位的驱动下,钙离子可由存在于线粒体内膜中的单向运送体输送进入线粒体基质;[46]排出线粒体基质时则需要钠-钙交换蛋白的辅助或通过钙诱导钙释放(calcium-induced-calcium-release,CICR)机制。[47]在钙离子释放时会引起伴随着较大膜电位变化的“钙波”(calcium wave),能激活某些第二信使系统蛋白,协调诸如突触中神经递质的释放及内分泌细胞中激素的分泌。线粒体也参与细胞凋亡时的钙离子信号转导。[48]
其他功能
除了合成ATP为细胞提供能量等主要功能外,线粒体还承担了许多其他生理功能。
- 调节膜电位并控制细胞程序性死亡:[49]当线粒体内膜与外膜接触位点处生成了由己糖激酶(细胞质基质蛋白)、外周苯并二氮受体和电压依赖阴离子通道(线粒体外膜蛋白)、肌酸激酶(线粒体膜间隙蛋白)、ADP-ATP载体(线粒体内膜蛋白)和亲环蛋白D(线粒体基质蛋白)等多种蛋白质组成的通透性转变孔道(PT孔道)后,会使线粒体内膜通透性提高,引起线粒体跨膜电位的耗散,从而导致细胞凋亡。[50]线粒体膜通透性增加也能使诱导凋亡因子(AIF)等分子释放进入细胞质基质,破坏细胞结构。[51]
- 细胞增殖与细胞代谢的调控;
- 合成胆固醇及某些血红素[52]。
线粒体的某些功能只有在特定的组织细胞中才能展现。例如,只有肝脏细胞中的线粒体才具有对氨气(蛋白质代谢过程中产生的废物)造成的毒害解毒的功能。[53]
起源假说
对于线粒体的起源有两种假说,分别为内共生假说与非内共生假说:
内共生假说
该假说认为线粒体起源于被另一个细胞吞噬的线粒体祖先——原线粒体——一种能进行三羧酸循环和电子传递的革兰氏阴性菌。这种好氧细菌是变形菌门下的一个分支,与立克次氏体有密切关系。原线粒体被吞噬后,并没有被消化,而是与宿主细胞形成了共生关系——寄主可以从宿主处获得更多营养,而宿主则可使用寄主产生的能量——这种关系增加了细胞的竞争力,使其可以适应更多的生存环境。在长期对寄主和宿主都有利的互利共生中,原线粒体逐渐演变形成了线粒体,使宿主细胞中进行的糖酵解和原线粒体中进行的三羧酸循环和氧化磷酸化成功耦合。[54]有研究认为,这种共生关系大约发生在17亿年以前,[55],与进化趋异产生真核生物和古细菌的时期几乎相同。[56]但线粒体与真核生物细胞核出现的先后关系仍存在争议。[57]
现已发现支持内共生学说的证据包括:
- 线粒体有內膜,拥有自己的DNA,其形状与细菌的环状DNA类似;
- 线粒体的DNA上编码了在线粒体中表达的特定蛋白质;
- 线粒体的遗传密码与变形菌门细菌的遗传密码更为相似[58];
- 线粒体核糖体不论在大小还是在结构上都与细菌70S核糖体较为相似,而与真核细胞的80S核糖体差异较大。[59]
非内共生假说
非内共生假说又称为“细胞分化学说”,认为线粒体的发生是由细胞膜或内质网膜等生物膜系统中的膜结构演变而来的。非内共生学说有几种模型,主流的模型认为在细胞进化的最初阶段,原核细胞基因组复制后并不伴有典型的无丝分裂,而是拟核附近的细胞膜内陷形成双层膜,将其中一个基因组包围、隔离,进而发生细胞分裂。未分裂出来的子细胞则缓慢演化为细胞核、线粒体和叶绿体等高度特化的细胞结构。
遗传学
基因组
线粒体的基因组中基因的数量很少,规模远小于细菌基因组。但内共生学说认为线粒体源于被吞噬的细菌,那么两者基因组规模应该较为相似。为了解释这一现象,有猜想认为原线粒体的基因除了丢失了一些外,大部分转移到了宿主细胞的细胞核中,[60]所以核基因编码了在超过98%的线粒体表达内的蛋白质。[61]某些线粒体中不含DNA的生物(如隐孢子虫等)的mtDNA可能已完全丢失或整合入核DNA中。[62]线粒体DNA(mtDNA)在线粒体中有2-10个备份,[63]呈双链环状(但也有呈线状的特例存在[64])。mtDNA长度一般为几万至数十万碱基对,人类mtDNA的长度为16,569bp,[65]拥有有37个基因,编码了两种rRNA(12S rRNA和16S rRNA)、22种tRNA(同样转运20种标准氨基酸,只是亮氨酸和丝氨酸都有两种对应的tRNA)以及13种多肽(呼吸链复合物Ⅰ、Ⅲ、Ⅳ、Ⅴ的亚基)。[66]mtDNA的长度和线粒体基因组的大小因物种而异,表一列出了几种模式生物mtDNA的长度:
| 生物 | 学名 | mtDNA长度(bp) |
|---|---|---|
芽殖酵母 | Saccharomyces cerevisiae | 85779[67] |
裂殖酵母 | Schizosaccharomyces pombe | 19431[68] |
拟南芥 | Arabidopsis thaliana | 366924[69] |
水稻 | Oryza sativa | 490520[70] |
秀丽隐杆线虫 | Caenorhabditis elegans | 13794[71] |
黑腹果蝇 | Drosophila melanogaster | 19517[72] |
非洲爪蟾 | Xenopus laevis | 17553[73] |
小鼠 | Mus musculus | 16300[74] |
mtDNA利用率极高,线粒体基因组各基因之间排列十分紧凑,部分区域还可能出现重叠(即前一个基因的最后一段碱基与下一个基因的第一段碱基相衔接)。人类mtDNA中基因间隔区总共只有87bp,占mtDNA总长的0.5%。[75]mtDNA的两条DNA单链均有编码功能,其中重链编码两个rRNA、12个mRNA和14个tRNA;轻链编码一个mRNA和8个tRNA。[76]mtDNA一般没有内含子(如人类的mtDNA等),[77]但也已发现某些真核生物的mtDNA拥有内含子,[78]这些生物包括:盘基网柄菌[79]等原生生物[80]和酵母菌(其OXi3基因有9个内含子)[81]。这些mtDNA中的内含子在基因转录产物的加工和翻译中可能有一定功能。[82]
线粒体基因组通常都是存在于同一个mtDNA分子中,但少数生物的线粒体基因组却分别储存在多个不同的mtDNA中。例如,人虱的线粒体基因组就分开储藏于18个长约3-4kb的微型环状DNA中,每个DNA分子只分配到了1-3个基因。[83]这些微型环状DNA之间也存在着同源或非同源的基因重组现象,但成因未知。[84]
遗传密码
线粒體中拥有一套独特的遗传系统。在进行人类線粒體遗传学研究时,人们确认線粒體的遗传密码与通用遗传密码也有些许差异。[85]自从上述发现证明并不只存在单独的一种遗传密码之后,许多有轻微不同的遗传密码都陆续被发现。[86]在線粒體的遗传密码中最常见的差异是:AUA由異白胺酸变为甲硫氨酸的密码子、UGA由终止密码子变为色氨酸的密码子、AGA和AGG由精氨酸的密码子变为终止密码子(植物等生物的线粒体遗传密码另有差异,参见表二)。[87]此外,也有某些特例是只涉及终止密码子的,在山羊支原体线粒体遗传密码的UGA由终止密码子变为色氨酸的密码子,而且使用频率比UGG更高;[88]四膜虫線粒體遗传密码里只有UGA一种终止密码子,其UAA和UAG由终止密码子变为谷氨酰胺的密码子;而游仆虫線粒體遗传密码里则只有UAA和UAG两种终止密码子,其UGA由终止密码子变为半胱氨酸的密码子。[89]通过線粒體遗传密码和通用遗传密码的对比,可以推导出遗传密码演化过程的可能模式。[90]
| 密码子 | 通用密码 | 線粒體遗传密码 | |||
真菌 | 植物 | 无脊椎动物 | 哺乳动物 | ||
| UGA | 终止密码子 | 色氨酸 | 终止密码子 | 色氨酸 | 色氨酸 |
| AUA | 异亮氨酸 | 甲硫氨酸 | 异亮氨酸 | 甲硫氨酸 | 甲硫氨酸 |
| CUA | 亮氨酸 | 苏氨酸 | 亮氨酸 | 亮氨酸 | 亮氨酸 |
| AGA、AGG | 精氨酸 | 精氨酸 | 精氨酸 | 丝氨酸 | 终止密码子 |
分裂与融合
线粒体的融合是与分裂协同进行的,过程高度保守,需要在多种蛋白质的精确调控下完成。[91]两者一般保持动态平衡,这种平衡对维持线粒体正常的形态、分布和功能十分重要。线粒体融合与分裂间的失衡可产生巨型线粒体,这种过大的线粒体常见于病变的肝细胞、恶性营养不良患者的胰脏细胞和白血病患者骨髓的巨噬细胞中。[92]分裂异常会导致线粒体破碎,而融合异常则会导致线粒体形态延长,两者都会影响线粒体的功能。[93]分裂与融合活动异常的线粒体膜电位通常会降低,并最终经线粒体自噬作用清除。
线粒体的分裂在真核细胞内经常发生。为了保证在细胞发生分裂后每个子细胞都能继承母细胞的线粒体,母细胞中的线粒体在一个细胞周期需要至少复制一次。即使是在不再分裂的细胞内,线粒体为了填补已老化的线粒体造成的空缺也需要进行分裂。[94]的线粒体以与细菌的无丝分裂类似的方式进行增殖,可细分为三种模式:[95]
- 间壁分离(见于部分动物和植物线粒体):线粒体内部首先由内膜形成隔,随后外膜的一部分内陷,插入到隔的双层膜之间,将线粒体一分为二。
- 收缩分离(见于蕨类植物和酵母菌线粒体):线粒体中部先缢缩同时向两端不断拉长然后一分为二。
- 出芽分离(见于藓类植物和酵母菌线粒体):线粒体上先出现小芽,小芽脱落后成长、发育为成熟线粒体。
线粒体的融合也是细胞中的基本事件,对线粒体正常功能的发挥具有非常重要的作用。人类细胞需要通过线粒体融合的互补作用来抵抗衰老;酵母细胞线粒体融合发生障碍会引起呼吸链缺陷。[96]线粒体间的融合需在一种分子量约为800kDa的蛋白质复合物——“融合装置”(fusion machinery)的介导下进行,[97]该过程可大致分为四个步骤:锚定、外膜融合、内膜融合以及基质内含物融合。[98]
群体遺傳學
因为mtDNA几乎不发生基因重组,所以遗传学家长期将其作为研究群体遗传学与进化生物学的信息来源。[99]所有mtDNA是以单一单元(单体型)进行遗传的(而不像细胞核中的DNA储存在多个染色体中),它们在亲本与子代之间的传递关系并不复杂,因此不同个体间mtDNA的联系便可以利用系统发生树来表现。[100]而从这些系统发生树的形态中人们可以得知种群的进化史。人类进化遗传学中运用分子钟技术推算出了线粒体夏娃最晚出现的时间[101](这个成果被认为是人类由非洲单地起源的有力依据[102])是利用mtDNA研究群体遗传学的典型例子。另外一个例子是对尼安德特人骨骼化石中mtDNA测序。该测序的结果显示,尼安德特人与解剖学意义上的现代人在mtDNA序列上有较大差异,说明两者间缺乏基因交流。[103]虽然mtDNA在遗传学研究中占据了重要地位,但是mtDNA序列中的信息只能反映所考察的群体中的雌性成员的演化进程,而不能代表整个种群。这一缺陷需要由对父系遗传序列(如Y染色体上的非重组区)的测序弥补。[104]广义上来说,只有既考虑了mtDNA又考虑了核DNA的遗传学研究才能为种群的进化史提供全面的线索。[105]
机能障碍与疾病
正常細胞含數個至千餘個相同的粒線體,如細菌大小。研究證實,在老人身上,其身體細胞內粒線體的含量有明顯減少。粒線體負責製造腺苷三磷酸ATP,如同發電機一般,是身體能量的來源,其在轉換為ATP能量的過程需動用電子傳遞。如果沒有正確捕捉到電子,逸出的電子會與氧分子結合成超氧自由基,很容易破壞鹼基而造成粒線體DNA突變,進而累積一些細胞的衰老或疾病因子,像是一些老年疾病:糖尿病、心臟病、關節炎等,都與粒線體DNA變異有關。
粒線體缺陷疾病
線粒體病(mitochondrial disorders)是遺傳缺損引起線粒體代謝酶缺陷,致使ATP合成障礙、能量來源不足導致的一組異質性病變。
線粒體是密切與能量代謝相關的細胞器,無論是細胞的存活(氧化磷酸化)和細胞死亡(凋亡)均與線粒體功能有關,特別是呼吸鏈的氧化磷酸化異常與許多人類疾病有關。
Luft等(1962)首次報道一例線粒體肌病,生化研究證實為氧化磷酸化脫耦聯引起。Anderson(1981)測定人類線粒體DNA(mtDNA)全長序列,Holt(1988)首次發現線粒體病患者mtDNA缺失,證實mtDNA突變是人類疾病的重要病因,建立了有別于傳統孟德爾遺傳的線粒體遺傳新概念。
根據線粒體病變部位不同可分為:
1.線粒體肌病(mitochondrial myopathy)線粒體病變侵犯骨骼肌為主。
2.線粒體腦肌病(mitochondrial encephalomyopathy)病變同時侵犯骨骼肌和中樞神經系統。
3.線粒體腦病病變侵犯中樞神經系統為主。
参见
- 內共生理論
- 呼吸作用
- 化学渗透假说
- 叶绿体
- 线粒体病
- 线粒体DNA
- 糖酵解
- 线粒体遗传学
- 线粒体核糖体
紡錘核移植(Spindle transfer/spindle nuclear transfer)
線粒體病(Mitochondrial disease)
注释
^ 这些现在已不再继续使用的名称包括:blepharoblast、condriokont、chondriomite、chondrioplast、chondriosome、chondrioshere、filum、fuchsinophilic granule、interstitial body、körner、fädenkörner、mitogel、parabasal body、plasmasome、plastochondria、plastome、sphereoplast和vermicle等(按首字母在英文字母表中的顺序排列),其中“chondriosome”(可译为“颗粒体”)直至1982年仍见诸欧洲各国的科学文献。
^ 现在人们已经了解前人观察细胞时发现线粒体呈现不同形状的原因:一方面,线粒体自身形状多变;另一方面,制备切片时切割细胞的角度不同也会影响到显微镜下观察到的线粒体的形状。
^ 线粒体基质中每1μL的水溶解了约1.25mg的蛋白质,而细胞质基质中每1μL的水中只溶解了约0.26mg蛋白质,所以线粒体基质较细胞质基质黏稠。
^ 有氧呼吸第一阶段对应的是糖酵解,是糖类经过一系列酶促反应产生丙酮酸的过程。该过程在细胞质基质内完成,能释放少量能量。
参考文献
^ 朱正威、赵占良等. 生物1分子与细胞. 北京市海淀区中关村南大街17号院1号楼: 人民教育出版社. 2007年: 53–54 [2011年]. ISBN 978-7-107-17670-8 (中文).
^ Henze K, Martin W. Evolutionary biology: essence of mitochondria. Nature. 2003-11-13, 426 (6963): 127–128. PMID 14614484. doi:10.1038/426127a (英语).
^ Campbell, Neil A.; Brad Williamson; Robin J. Heyden. Biology: Exploring Life. Boston, Massachusetts: Pearson Prentice Hall. 2006. ISBN 0-13-250882-6 (英语). 引文使用过时参数coauthors (帮助)
^ McBride HM, Neuspiel M, Wasiak S. Mitochondria: more than just a powerhouse. Curr. Biol. Jul 25, 2006, 16 (14): R551–R560. PMID 16860735. doi:10.1016/j.cub.2006.06.054 (英语).
^ William Bechtel. Discovering cell mechanisms: the creation of modern cell biology. Cambridge University Press. Apr 28, 2008: 81. ISBN 978-0-521-72944-4 (英语).
^ 肖义军、俞如旺. 用高倍镜观察线粒体实验的建议. 生物学教学. 2011, (2): 60–60 (中文).
^ Bereiter‐Hahn J, Vöth M. Metabolic control of shape and structure of mitochondria in situ. Biology of the Cell. 1983, 47: 309–322 (英语).
^ Pieter Vanden Berghe, Grant W. Hennig, and Terence K. Smith. Characteristics of intermittent mitochondrial transport in guinea pig enteric nerve fibers. Am J Physiol Gastrointest Liver Physiol. Oct 30, 2003, 286 (4): G671–G682. doi:10.1152/ajpgi.00283.2003. (原始内容存档于2016年3月6日) (英语).
^ K. Van Gestel and J‐P. Verbelen. Giant mitochondria are a response to low oxygen pressure in cells of tobacco (Nicotiana tabacum L.). J. Exp. Bot. Jan 18, 2002, 53 (371): 1215–1218. doi:10.1093/jexbot/53.371.1215 (英语).
^ Robert E Jensen. Control of Mitochondrial Shape. Current Opinion in Cell Biology. Aug 2005, 17 (4): 384–388. doi:10.1016/j.ceb.2005.06.011 (英语).
^ Julie St-Pierre. Master of Mitochondrial Number (PDF). J Exp Biol 208, vii. Feb 1, 2005. doi:10.1242/jeb.01449 (英语).
^ 李依新、潘桂珍. 哺乳动物成熟红细胞的呼吸方式 (PDF). 生物学教学. 2007, 32 (8): 68–69 (中文). [永久失效連結]
^ 党永明、房亚东、胡炯宇等. 成体大鼠心肌细胞微管解聚对线粒体分布及能量代谢的影响. 中华烧伤杂志. 2010年二月, 26 (1). (原始内容存档于2011-12-15) (中文). 请检查|date=中的日期值 (帮助)
^ 刘姗、李媛、高选、杨慧军、颜军昊、陈子江. 人卵母细胞体外成熟前后线粒体分布的变化. 解剖学报. 2007, 38 (5) (中文).
^ 曲杨燕、刘云海、郭勇、李凯、赵雁伟、李晓燕、倪和民. 绵羊卵母细胞体外成熟过程中的线粒体分布变化 (PDF). 畜牧兽医学报. 2009, (8): 1258–1261 (中文).
^ Sidney Fleischer, George Rouser, Becca Fleischer, Anna Casu and Gene Kritchevsky. Lipid composition of mitochondria from bovine heart, liver, and kidney (PDF). The Journal of Lipid Research. May 1967, 8 (3): 170–180. PMID 4292227 (英语).
^ Albert von Kölliker (1817-1905) Würzburger histologist. Journal of the American Medical Association. Nov 25, 1968, 206 (9): 2111–2112. PMID 4880509. doi:10.1001/jama.1968.03150090187031 (英语).
^ Richard Altmann. Die elementarorganismen und ihre beziehungen zu den zellen. Veit. 1894. ISBN 978-3-8364-0891-2. OCLC 11276123 (德语).
^ Carl Benda. Weitere Mitteilungen über die Mitochondria. Verh Physiol Ges Berlin Jahrg. 1899, 99 (4-7): 376–383 (德语).
^ Leonor Michaelis. Die vitale Färbung, eine Darstellungsmethode der Zellgranula (PDF). Arch. Mikrosk. Anat. 1900, 55: 558–575 (德语). [永久失效連結]
^ Leonor Michaelis (with additions by D. A. Macinnes and S. Granick). Leonor Michaelis (PDF). 1958: 284 (英语).
^ Otto Heinrich Warburg. Über die Rolle des Eisens in der Atmung des Seeigeleis nebst Bemerkungen über einige durch Eisen beschleunigte Oxydationen. Hoppe-Seyler´s Zeitschrift für physiologische Chemie. Jan 1, 1914, 92 (3). ISSN 0018-4888. doi:10.1515/bchm2.1914.92.3.231 (德语).
^ David Keilin. On cytochrome, a respiratory pigment, common to animals, yeast, and higher plants. Proceedings of the Royal Society of London B: Biological Science. Aug 1, 1925, 98 (690): 312–339. doi:10.1098/rspb.1925.0039 (英语).
^ The Nobel Prize in Physiology or Medicine 1931. [2011-08-27].Your discovery about the nature and effect of the ferment of respiration, which the Caroline Institute is rewarding this year with Alfred Nobel's Prize for Physiology or Medicine, has added a link of brilliant achievement to the chain that binds for all time, John Mayow, Antoine Laurent Lavoisier, and Otto Warburg.
^ Herrmann JM, Neupert W. Protein transport into mitochondria. Current Opinion in Microbiology. Apr 4, 2000, 3 (2): 210–214. PMID 10744987. doi:10.1016/S1369-5274(00)00077-1 (英语).
^ Chipuk JE, Bouchier-Hayes L, Green DR. Mitochondrial outer membrane permeabilization during apoptosis: the innocent bystander scenario. Cell Death and Differentiation. May 19, 2006, 13 (8): 1396–1402. PMID 16710362. doi:10.1038/sj.cdd.4401963 (英语).
^ J D Robertson, B Zhivotovsky, V Gogvadze, and S Orrenius. Outer mitochondrial membrane permeabilization: an open-and-shut case?. Cell Death and Differentiation. 2003, 10 (11): 485–487. doi:10.1038/sj.cdd.4401218 (英语).
^ PACS-2可控制内质网与线粒体之间的信息及介导Bid凋亡. 生命的化学. 2005, 25 (2): 168–168 (中文).
^ Hayashi T, Rizzuto R, Hajnoczky G, Su TP. MAM: more than just a housekeeper. Trends Cell Biol. Feb 2009, 19 (2): 81–88. PMC 2750097. PMID 19144519. doi:10.1016/j.tcb.2008.12.002 (英语).
^ 付玉龙、杨晓明. 线粒体膜间隙蛋白的转运机制. 医学分子生物学杂志. 2006, 3 (3) (中文).
^ McMillin JB, Dowhan W. Cardiolipin and apoptosis. Biochim Biophys Acta. Dec 30, 2002, 1585 (2-3): 97–107. PMID 12531542. doi:10.1016/S1388-1981(02)00329-3 (英语).
^ Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. 14. Molecular Biology of the Cell. New York: Garland Publishing Inc. 2002. ISBN 0-8153-3218-1 (英语). 引文使用过时参数coauthors (帮助)
^ Hanaki M, Tanaka K, Kashima Y. Scanning electron microscopic study on mitochondrial cristae in the rat adrenal cortex. Journal of Electron Microscopy. 1985, 34 (4): 373–380. PMID 3837809 (英语).
^ Frederick P. Prince. Mitochondrial cristae diversity in human Leydig cells: a revised look at cristae morphology in these steroid-producing cells. Anat Rec. Apr 1, 1999, 254 (4): 534–541. PMID 10203260 (英语).
^ Frederick P. Prince. Lamellar and tubular associations of the mitochondrial cristae: unique forms of the cristae present in steroid-producing cells. Mitochondrion. Feb 2002, 1 (4): 381–389. doi:10.1016/S1567-7249(01)00038-1 (英语).
^ Soboll, S., Scholz, R., Freisl, M., Elbers, R. & Heldt, H. W. Distribution of metabolites between mitochondria and cytosol of perfused liver. (编) J. M. Tager, Hans-Dieter Söling, John R. Williamson. Use of Isolated Liver Cells and Kidney Tubules in Metabolic Studies. New York: Elsevier. 1976: 29–40. ISBN 978-0444109255. OCLC 2137029. LCCN 76-009093 (英语).
^ 谢红卫、陈光辉、李绍清、谭艳平. 线粒体RNA编辑的研究进展. 作物研究. 2005, (5): 404–408 (英语).
^ O'Brien, T.W. The General Occurrence of 55S Ribosomes in Mammalian Liver Mitochondria (PDF). J. Biol. Chem. May 25, 1971, 245 (10): 3409–3417. PMID 4930061 (英语).
^ Jianzhong Jiang, Ying Zhang, Adrian R Krainer, Rui-Ming Xu. Crystal structure of human p32, a doughnut-shaped acidic mitochondrial matrix protein (PDF). Proceedings of the National Academy of Sciencesofthe United States of America. Mar 30, 1999, 96 (7): 3572–3577. PMID 10097078 (英语).
^ Reginald Garrett,Charles M. Grisham. 20. Electron Transport and Oxidative Phosphorylation. Biochemistry 4th. Mary Finch. Dec 2008: 623. ISBN 0-495-10935-5 (英语).
^ King A, Selak MA, Gottlieb E. Succinate dehydrogenase and fumarate hydratase: linking mitochondrial dysfunction and cancer. Oncogene. Aug 7, 2006, 25 (34): 4675–4682. PMID 16892081. doi:10.1038/sj.onc.1209594 (英语).
^ Huang H, Manton KG. The role of oxidative damage in mitochondria during aging: A review. Frontiers in Bioscience. May 1, 2004, 9: 1100–1117. PMID 14977532. doi:10.2741/1298 (英语).
^ Pizzo P, Pozzan T. Mitochondria–endoplasmic reticulum choreography: structure and signaling dynamics. Trends in Cell Biology. Oct 2007, 17 (10): 511–517. PMID 17851078. doi:10.1016/j.tcb.2007.07.011 (英语).
^ Editor-in-chief, George J. Siegel; editors, Bernard W. Agranoff; 等. 23. (编) Siegel GJ, Agranoff BW, Fisher SK, Albers RW, Uhler MD. Basic Neurochemistry 6. Lippincott Williams & Wilkins. 1999. part 3. ISBN 0-397-51820-X (英语). 引文格式1维护:显式使用等标签 (link)
^ CT Brighton, RM Hunt. Mitochondrial calcium and its role in calcification. Clinical Orthopaedics and Related Research. 1974, 100 (100): 406–416. PMID 4134194. doi:10.1097/00003086-197405000-00057 (英语).
^ Miller RJ. Mitochondria – the kraken wakes!. Trends in Neuroscience. Mar 1998, 21 (3): 95–97. PMID 9530913. doi:10.1016/S0166-2236(97)01206-X (英语).
^ 杨勇骥、汤莹、宋田斌、吴越、邰艳红、沙继宏、叶煦亭、郑尊. 骨骼肌兴奋收缩偶联时钙诱导钙释放机理的研究. 电子显微学报. 2000, 19 (3): 365–366 (中文).
^ Hajnóczky G, Csordás G, Das S, Garcia-Perez C, Saotome M, Sinha Roy S, Yi M. Mitochondrial calcium signalling and cell death: approaches for assessing the role of mitochondrial Ca2+ uptake in apoptosis. Cell Calcium. Nov–Dec 2006, 40 (5-6): 553–560. PMC 2692319. PMID 17074387. doi:10.1016/j.ceca.2006.08.016 (英语).
^ Green DR. Apoptotic pathways: the roads to ruin. Cell. Sep 18, 1998, 94 (6): 695–698. PMID 9753316. doi:10.1016/S0092-8674(00)81728-6 (英语).
^ Zamzami N, Hirsch T, Dallaporta B, Petit PX, Kroemer G. Mitochondrial implication in accidental and programmed cell death: apoptosis and necrosis (PDF). Journal of Bioenergetics and Biomembranes. Apr 2, 1997, 29 (2): 185–193. PMID 9239543. doi:10.1023/A:1022694131572 (英语).
^ 白世平、罗绪刚、吕 林. 线粒体在细胞凋亡中的介导作用. 生命科学. Aug 2006, 18 (4): 368–372 (中文). [永久失效連結]
^ Oh-hama T. Evolutionary consideration on 5-aminolevulinate synthase in nature. Origins of Life and Evolution of Biospheres. Aug 1997, 27 (4): 405–412. PMID 9249985. doi:10.1023/A:1006583601341 (英语).
^ Campbell JW, Smith DD Jr, Vorhaben JE. Avian and Mammalian Mitochondrial Ammonia-Detoxifying Systems in Tortoise Liver. Science. Apr 19, 1985, 228 (4697): 349–351. PMID 17790238. doi:10.1126/science.228.4697.349 (英语).
^ Emelyanov VV. Mitochondrial connection to the origin of the eukaryotic cell (PDF). European Journal of Biochemistry. Apr 2003, 270 (8): 1599–1618. PMID 12694174. doi:10.1046/j.1432-1033.2003.03499.x (英语).
^ Emelyanov VV. Rickettsiaceae, rickettsia-like endosymbionts, and the origin of mitochondria (PDF). Bioscience Reports. Feb 2001, 21 (1): 1–17. PMID 11508688. doi:10.1023/A:1010409415723 (英语).
^ Feng D-F, Cho G, Doolittle RF. Determining divergence times with a protein clock: update and reevaluation. Proceedings of the National Academy of Sciences of the United States of America. Nov 25, 1997, 94 (24): 13028–13033. PMC 24257. PMID 9371794. doi:10.1073/pnas.94.24.13028 (英语).
^ Gray MW, Burger G, Lang BF. Mitochondrial evolution. Science (journal). Mar 5, 1999, 283 (5407): 1476–1481. PMID 10066161. doi:10.1126/science.283.5407.1476 (英语).
^ Futuyma DJ. On Darwin's Shoulders. Natural History. Nov 1, 2005, 114 (9): 64–68 (英语).
^ O'Brien TW. Properties of human mitochondrial ribosomes. IUBMB Life. Sep 2003, 55 (9): 505–513. PMID 14658756. doi:10.1080/15216540310001626610 (英语).
^ Ryan MT, Hoogenraad NJ. Mitochondrial-nuclear communications (PDF). Annual Review of Biochemistry. 2007, 76: 701–722. ISSN 0066-4154. PMID 17227225. doi:10.1146/annurev.biochem.76.052305.091720 (英语). [永久失效連結]
^ Stolzenberg N, Nguyen The B, Salducci MD, Cavalli L. Influence of Environment and Mitochondrial Heritage on the Ecological Characteristics of Fish in a Hybrid Zone. PLoS One. Jun 18, 2009, 4 (6): e5962. PMID 19536286. doi:10.1371/journal.pone.0005962 (英语).
^ Henriquez FL, Richards TA, Roberts F, McLeod R, Roberts CW. The unusual mitochondrial compartment of Cryptosporidium parvum. Trends in Parasitology. 2005 February, 21 (2): 68–74. PMID 15664529. doi:10.1016/j.pt.2004.11.010 (英语). 请检查|date=中的日期值 (帮助)
^ Wiesner RJ, Ruegg JC, Morano I. Counting target molecules by exponential polymerase chain reaction, copy number of mitochondrial DNA in rat tissues. Biochim Biophys Acta. Mar 16, 1992, 183 (2): 553–559. PMID 1550563 (英语).
^ Fukuhara H, Sor F, Drissi R, Dinouël N, Miyakawa I, Rousset, and Viola AM. Linear mitochondrial DNAs of yeasts: frequency of occurrence and general features (PDF). Molecular and Cellular Biology. Apr 1993, 13 (4): 2309–2314. PMC 359551. PMID 8455612 (英语).
^ Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR; 等. Sequence and organization of the human mitochondrial genome. Nature. Apr 9, 1981, 410 (5806): 141. PMID 7219534. doi:10.1038/290457a0 (英语). 引文格式1维护:显式使用等标签 (link)
^ Chan DC. Mitochondria: Dynamic Organelles in Disease, Aging, and Development. Cell. Jun 30, 2006, 125 (7): 1241–1252. PMID 16814712. doi:10.1016/j.cell.2006.06.010 (英语).
^ Foury F, Roganti T, Lecrenier N, Purnelle B. The complete sequence of the mitochondrial genome of Saccharomyces cerevisiae (PDF). FEBS Letters. Dec 4, 1998, 440 (3): 325–331. PMID 9872396. (原始内容 (PDF)存档于2007-07-20) (英语).
^ BF Lang, R Cedergren, MW Gray. The mitochondrial genome of the fission yeast Schizosaccharomyces pombe: highly homologous introns are inserted at the same position of the otherwise less conserved cox1 genes in Schizosaccharomyces pombe and Aspergillus nidulans. European Journal of Biochemistry. Sep 1984, 3 (9): 2129–2136. PMC 557654. PMID 6092057. doi:10.1111/j.1432-1033.1987.tb13641.x (英语).
^ J Marienfeld, M Unseld, P Brandt, A Brennicke. Genomic recombination of the mitochondrial atp6 gene in Arabidopsis thaliana at the protein processing site creates two different presequences (PDF). DNA Research. 1996, 3 (5): 287–290. PMID 9039497. doi:10.1093/dnares/3.5.287 (英语).
^ Y. Notsu, S. Masood, T. Nishikawa, N. Kubo, G. Akiduki, M. Nakazono, A. Hirai and K. Kadowaki. The complete sequence of the rice (Oryza sativa L.) mitochondrial genome: frequent DNA sequence acquisition and loss during the evolution of flowering plants (PDF). Molecular Genetics and Genomics. Nov 4, 2002, 268 (4): 434–445. PMID 12471441. doi:10.1007/s00438-002-0767-1 (英语).
^ R Okimoto, JL Macfarlane, DR Wolstenholme. Evidence for the frequent use of TTG as the translation initiation codon of mitochondrial protein genes in the nematodes, Ascaris suum and Caenorhabditis elegans. Nucleic Acids Research. Oct 25, 1990, 18 (20): 6113–6118. PMC 332414. PMID 2235493. doi:10.1093/nar/18.20.6113 (英语).
^ Douglas O. Clary, Judy M. Goddard*, Susan C. Martin, Christiane M.-R. Fauron† and David R. Wolstenholme. Drosophila mitochondrial DNA: a novel gene order (PDF). Nucleic Acids Research. Nov 11, 1982, 10 (21): 6619–6637. PMC 326953. PMID 6294611. doi:10.1093/nar/10.21.6619 (英语).
^ Wong JF, Ma DP, Wilson RK, Roe BA. DNA sequence of the Xenopus laevis mitochondrial heavy and light strand replication origins and flanking tRNA genes. Nucleic Acids Res. Jul 25, 1983, 11 (14): 4977–4995. PMC 326100. PMID 6308566 (英语).
^ Goios A, Pereira L, Bogue M, Macaulay V, Amorim A. mtDNA phylogeny and evolution of laboratory mouse strains (PDF). Genome Research. Feb 6, 2007, 17 (3): 293–298. PMC 1800920. PMID 17284675. doi:10.1101/gr.5941007 (英语).
^ РИ Сукерник, ОА Дербенева, ЕБ Стариковская, НВ Володько, ИЕ Михайловская, ИЮ Бычков, МТ Лотт, МД Браун, ДК Уоллес. Митохондриальный геном и митохондриальные болезни человека [The Mitochondrial Genome and Human Mitochondrial Diseases] (PDF). Генетика. 2002, 38 (2): 1–10. PMID 11898607. doi:10.1023/A:1014369624346 (俄语).
^ DA Clayton. Transcription of the Mammalian Mitochondrial Genome. Annual Review of Biochemistry. 1984, 53: 573–594. PMID 6383200. doi:10.1146/annurev.bi.53.070184.003041 (英语).
^ Anderson S, Bankier AT, Barrell BG, de-Bruijn MHL, Coulson AR; 等. Sequence and organization of the human mitochondrial genome. Nature. Apr 9, 1981, 290 (5806): 427–465. PMID 7219534. doi:10.1038/290457a0 (英语). 引文格式1维护:显式使用等标签 (link)
^ Bernardi G. Intervening sequences in the mitochondrial genome. Nature. Dec 7, 1978, 276 (5688): 558–559. PMID 214710. doi:10.1038/276558a0 (英语).
^ Gray MW, Lang BF, Burger G. Mitochondria of protists. Annual Review of Genetics. 2004, 38: 477–524. PMID 15568984. doi:10.1146/annurev.genet.37.110801.142526.
^ Gray MW, Lang BF, Cedergren R, Golding GB, Lemieux C, Sankoff D; 等. Genome structure and gene content in protist mitochondrial DNAs (PDF). Nucleic Acids Research. 1998, 26 (4): 865–878. PMC 147373. PMID 9461442. doi:10.1093/nar/26.4.865 (英语). 引文格式1维护:显式使用等标签 (link)
^ Hebbar SK, Belcher SM, Perlman PS. A maturase-encoding group IIA intron of yeast mitochondria self-splices in vitro (PDF). Nucleic Acids Research. Apr 1992, 20 (7): 1747–1754. PMC 312266. PMID 1579468. doi:10.1093/nar/20.7.1747 (英语).
^ 齐义鹏. 酵母线粒体细胞色素b基因突变的定位和内含子的功能. 科学通报. 1986, 17: 1342–1346 (中文). [永久失效連結]
^ Shao R, Kirkness EF, Barker SC. The single mitochondrial chromosome typical of animals has evolved into 18 minichromosomes in the human body louse, Pediculus humanus. Genome Research. Mar 2009, 19 (5): 904–912. PMC 2675979. PMID 19336451. doi:10.1101/gr.083188.108 (英语).
^ Shao R, Barker SC. Chimeric mitochondrial minichromosomes of the human body louse, Pediculus humanus: evidence for homologous and non-homologous recombination. Gene. Feb 15, 2011, 473 (1): 36–43. ISSN 0378-1119. PMID 21092752. doi:10.1016/j.gene.2010.11.002 (英语).
^ Barrell BG, Bankier AT, Drouin J. A different genetic code in human mitochondria. Nature. Nov 8, 1979, 282 (5735): 189–194. PMID 226894. doi:10.1038/282189a0 (英语).
^ Jukes TH, Osawa S. The genetic code in mitochondria and chloroplasts. Experientia. Dec 1, 1990, 46 (11-12): 1117–1126. PMID 2253709. doi:10.1007/BF01936921 (英语).
^ SM Khan, RM Smigrodzki, R Swerdlow. Cell and animal models of mtDNA biology: progress and prospects (PDF). American Journal of Physiology-Cell Physiology. Feb 2007, 292 (2): C658–C669. PMID 16899549. doi:10.1152/ajpcell.00224.2006 (英语).
^ F Yamao, A Muto, Y Kawauchi, M Iwami, S Iwagami, Y Azumi, and S Osawa. UGA is read as tryptophan in Mycoplasma capricolum (PDF). Proceedings of the National Academy of Sciences. Apr 1985, 82 (8): 2306–2309. PMC 397546. PMID 3887399. doi:10.1073/pnas.82.8.2306 (英语).
^ Salas-Marco J, Fan-Minogue H, Kallmeyer AK, Klobutcher LA, Farabaugh PJ, Bedwell DM. Distinct paths to stop codon reassignment by the variant-code organisms Tetrahymena and Euplotes (PDF). Molecular and Cellular Biology. Jan 2006, 26 (2): 438–447. PMC 1346903. PMID 16382136. doi:10.1128/MCB.26.2.438–447.2006 请检查|doi=值 (帮助) (英语).
^ 陈惟昌、陈志华、王自强、刘伟、左琳. 線粒體遗传密码及基因组遗传密码的对称分析. 生物物理学报. Mar 2002, 18 (1): 87–94 (英语).
^ 蒋春笋、肖伟明、陈佺. 线粒体分裂、融合与细胞凋亡. 生物物理学报. 2007, 23 (4): 256–264 (中文).
^ Hermann GJ, Thatcher JW, Mills JP, Hales KG, Fuller MT, Nunnari J, Shaw JM. Mitochondrial Fusion in Yeast Requires the Transmembrane GTPase Fzo1p (PDF). the Journal of Cell Biology. Oct 19, 1998, 143 (2): 359–373. PMC 2132826. PMID 9786948. doi:10.1083/jcb.143.2.359 (英语).
^ Diaz F, Moraes CT. Mitochondrial biogenesis and turnover. Cell Calcium. Jul 2008, 44 (1): 24–35. PMID 18395251 (英语).
^ Alberts B, Johnson A, Lewis J; 等. 14. Energy Conversion: Mitochondria and Chloroplasts (The Genetic Systems of Mitochondria and Plastids). Molecular Biology of the Cell 4th. New York: Garland Science. 2002. ISBN 0-8153-3218-1 (英语). 引文格式1维护:显式使用等标签 (link)
^ 孟紫强、耿红. 与线粒体分裂有关的蛋白质研究进展 (PDF). 生命的化学. 2002, 22 (2): 118–120 (中文). [永久失效連結]
^ Westermann B. Merging mitochondria matters: cellular role and molecular machinery of mitochondrial fusion. EMBO Reports. Jun 2002, 3 (6): 527–531. ISSN 1469-221X. PMC 1084147. PMID 12052774. doi:10.1093/embo-reports/kvf113 (英语).
^ Westermann B. Molecular machinery of mitochondrial fusion and fission (PDF). Journal of Biological Chemistry. Mar 27, 2008, 283 (20): 13501–13505. PMID 18372247. doi:10.1074/jbc.R800011200 (英语).
^ 耿红、孟紫强. 线粒体融合机制研究进展. 细胞生物学杂志. 2003, 25 (1): 17–21 (中文).
^ RamonCastro JA, Picornell A, Ramon M. Mitochondrial DNA: a tool for populational genetics studies (PDF). International Microbiology. Dec 1998, 1 (4): 327–332. PMID 10943382 (英语).
^ Torroni A, Achilli A, Macaulay V, Richards M, Bandelt HJ. Harvesting the fruit of the human mtDNA tree. Trends Genet. Jun 2006, 22 (6): 339–345. PMID 16678300. doi:10.1016/j.tig.2006.04.001 (英语).
^ Cann RL, Stoneking M, Wilson AC. Mitochondrial DNA and human evolution (PDF). Nature. Jan 1, 1987, 325 (6099): 31–36. PMID 3025745. doi:10.1038/325031a0 (英语). [永久失效連結]
^ Garrigan D, Hammer MF. Reconstructing human origins in the genomic era. Nature Reviews Genetics. Sep 2006, 7 (9): 669–680. PMID 16921345. doi:10.1038/nrg1941 (英语).
^ Krings M, Stone A, Schmitz RW, Krainitzki H, Stoneking M, Pääbo S. Neandertal DNA sequences and the origin of modern humans (PDF). Cell. Jul 11, 1997, 90 (1): 19–30. PMID 9230299. doi:10.1016/S0092-8674(00)80310-4. (原始内容 (PDF)存档于2011-03-04) (英语).
^ Peter A. Underhill, Peidong Shen, Alice A. Lin, Li Jin, Giuseppe Passarino, Wei H. Yang, Erin Kauffman, Batsheva Bonné-Tamir, Jaume Bertranpetit, Paolo Francalacci, Muntaser Ibrahim, Trefor Jenkins, Judith R. Kidd, S. Qasim Mehdi, Mark T. Seielstad, R. Spencer Wells, Alberto Piazza, Ronald W. Davis, Marcus W. Feldman, L. Luca Cavalli-Sforza & Peter. J. Oefner. Y chromosome sequence variation and the history of human populations. Nature Genetics. Nov 2000, 26 (3): 358–361. PMID 11062480. doi:10.1038/81685 (英语).
^ Harding RM, Fullerton SM, Griffiths RC, Bond J, Cox MJ, Schneider JA, Moulin DS, Clegg JB. Archaic African and Asian lineages in the genetic ancestry of modern humans (PDF). American Journal of Human Genetics. Apr 1997, 60 (4): 772–789. PMC 1712470. PMID 9106523 (英语).
外部連結
线粒体研究(mitochondrial.net)
线粒体:功能决定结构(cytochemistry.net)- MIP,线粒体生理学学会
密西根大学提供的结构研究:
- 线粒体外膜蛋白的3维结构
- 线粒体内膜蛋白的3维结构
阿拉巴马大学提供的线粒体相关链接
美因茨大学提供的线粒体图册
威斯康星大学提供的线粒体蛋白间的合作关系研究资料
- Cell Centered Database中的线粒体资料
圣地亚哥州立大学提供的线粒体电子断层扫描重建资料
- 大鼠肝脏线粒体的低温电子断层扫描视频
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
|

