细菌
| 细菌界 化石时期:太古宙或更早 – 現今 冥古宙 太古宙 元古宙 显 | |
|---|---|
 | |
掃描式電子顯微鏡(SEM)下的大腸桿菌。 | |
科学分类 | |
| 域: | 细菌域 Bacteria |
| 界: | 细菌界 Bacteria |
| 門[1] | |
放線菌門 Actinobacteria(高G+C)
產水菌門 Aquificae
酸桿菌門 Acidobacteria | |

革蘭氏陽性菌的結構圖
細菌(学名:Bacteria)是生物的主要類群之一,屬於細菌域。也是所有生物中數量最多的一類,據估計,其總數約有5×1030個[2]。细菌是非常古老的生物,大约出现于37亿年前。真核生物细胞中的两种细胞器:线粒体和叶绿体,通常被认为是来源于内共生细菌。微生物无处不在,只要是有生命的地方,都会有微生物的存在。它们存在于人类呼吸的空气中,喝的水中,吃的食物中。细菌可以被气流从一个地方带到另一个地方。人体是大量细菌的栖息地;可以在皮肤表面、肠道、口腔、鼻子和其他身体部位找到。
細菌的個體非常小,目前已知最小的細菌只有0.2微米長[3],因此大多只能在顯微鏡下看到它們;而世界上最大的細菌可以用肉眼直接看見,有0.2-0.6毫米大,是一種叫納米比亞嗜硫珠菌的細菌。細菌一般是單細胞,細胞結構簡單,缺乏細胞核以及膜狀胞器,例如粒線體和葉綠體。基於這些特徵,細菌屬於原核生物。原核生物中還有另一類生物稱做古細菌,是科學家依據演化關係而另闢的類別。為了區別,本類生物也被稱做真細菌(Eubacteria)。古細菌與真細菌在生活環境、營養方式以及遺傳上有所不同。細菌的形狀相當多樣,主要有球狀、桿狀,以及螺旋狀。
細菌廣泛分佈於土壤和水中,或著與其他生物共生。人體身上也帶有相當多的細菌。據估計,人體內及表皮上的細菌細胞總數約是人體細胞總數的十倍[4]。此外,也有部分種類分布在極端的環境中,例如溫泉,甚至是放射性廢棄物中[5],它們被歸類為嗜極生物,其中最著名的種類之一是海棲熱袍菌,科學家是在意大利的一座海底火山中發現這種細菌的[6]。甚至在太空梭上也能生長[7]。然而,細菌種類是如此多,科學家研究過並命名的種類只佔其中的小部份。細菌域下所有門中,只有約一半能在實驗室培養的種類[8]。
細菌的營養方式有自养及异养,其中异养的腐生細菌是生态系统中重要的分解者,使碳循環能順利進行。部分細菌會進行固氮作用,使氮元素得以轉換為生物能利用的形式。細菌也對人類活動有很大的影響。一方面,細菌是許多疾病的病原體,包括肺結核、淋病、炭疽病、梅毒、鼠疫、砂眼等疾病都是由細菌所引發。然而,人類也時常利用細菌,例如乳酪及酸奶和酒釀的製作、部分抗生素的製造、廢水的處理等,都與細菌有關。在生物科技領域中,細菌有也著廣泛的運用。
在一般的情況下,一公克的土壤會大約含有4000萬隻細菌,一毫升的純水則約含有100萬隻細菌。[來源請求]總的來說,這世界上約有5×1030 隻細菌[9]。其生物量遠大於世界上所有動植物體內細胞數量的總和。[10]細菌還在營養素循環上扮演相當重要的角色,像是微生物造成的腐敗作用,就與氮循環相關。而在海底火山和在冷泉中,細菌則是靠硫化氫和甲烷來產生能量。2013年3月17日,研究者在深約11公里的馬里亞納海溝中發現了細菌。[11][12]其他研究則指出,在美國西北邊離岸2600米的海床下580米深處,仍有許多的微生物[11][13]根據這些研究人員的說法:「你可以在任何地方找到他們,他們的適應力遠比你想像的還要強,可以在任何地方存活。」[11]
目录
1 演化學
2 形態學
2.1 分类地位
3 結構
4 繁殖
5 代谢
6 运动
7 成長與繁殖
8 個體分裂
9 遺傳學
10 行為
10.1 细菌分类
11 與其他生物的交互關係
11.1 用處和危害
12 历史
13 参见
14 參考文獻
14.1 引用
14.2 书籍
15 外部連結
演化學
現今的細菌是從40億年前的單細胞生物演化而來。在此後的30億年間,細菌和古細菌都是主要的生物[14][15]。雖然細菌有化石存在,如疊層石等,但這些化石缺乏有效的形態學證據,很難與現生的細菌共同建構出細菌的演化史。幸運的是,日益成熟的基因定序技術讓我們有機會建立演化的樹狀圖,這些研究使我們明瞭了細菌演化的第一次大分歧是在真核及原核之間。[16]
之後,細菌又發生了第二次的劇烈演化,有一部分的古細菌與其他細菌內共生,成為了現今真核生物的祖先[17][18]。有部分科學家提出假說,認為真核生物的祖先吞下了一種α-變形菌門的細菌,成為後來的粒線體,或是氫酶體。之後,有些已經擁有粒線體的生物,吞下了類似藍菌類的生物,形成了後來的葉綠體,這一支後來演化成了藻類和植物。另外,有些藻類還有可能再吞入其他藻類進行內共生,此假說稱為二次內共生[19][20]。
形態學

細菌的形態學

在俄勒冈州米奇温泉流出的嗜热细菌生物薄膜,约20毫米厚。
細菌無論是在體型上或型態上都具有相當巨大的變異,有關細菌型態上的學門即被稱為型態學。
細菌細胞的大小大約是真核生物細胞的十分之一,大約 0.5–5.0 微米長。然而,有一些種類(如纳米比亚嗜硫珠菌)可以達半毫米長,甚至用肉眼就可以辨識。[21][22]。最小的細菌則是黴漿菌,長度大概只有 0.3 μm,與最大的病毒差不多大。[23]有些細菌甚至更小,但這些超微細菌還沒有完全被研究,超微細菌並不是一個演化上的分類,而是對於這些細微細菌的通稱。[24]
細菌通常以外觀分為球狀的球菌(coccus)、棒狀的桿菌(bacilli),游泳時個體會有所延展[25]。另外還有逗點狀的弧菌(vibrio),和螺旋體(spirilla),還有一小部分的細菌是二十面體或是立方體[26] 最近,在地殼深處發現了細菌,這種細菌生長為具有星形橫截面的分枝絲狀類型。這種大表面積與體積比的形態可以使這些細菌在營養貧乏的環境中具有優勢。[27]。
分类地位
细菌的分类的变化根本上反应了发展史思想的变化,许多种类甚至经常改变或改名。最近随着DNA测序,基因组学,生物信息学和计算生物学的发展,细菌学被放到了一个合适的位置。
最初除了蓝细菌外(它完全没有被归为细菌,而是归为蓝绿藻),其他细菌被认为是一类真菌。随着它们的特殊的原核细胞结构被发现,这明显不同于其他生物(它们都是真核生物),导致细菌归为一个单独的种类,在不同时期被称为原核生物,细菌,原核生物界。一般认为真核生物来源于原核生物。
通過研究rRNA序列,美國微生物學家卡尔·乌斯於1976年提出,原核生物包含两个大的類群。他将其称为真细菌(Eubacteria)和古细菌(Archaebacteria),后来被改名为细菌(Bacteria)和古菌(Archaea)。乌斯指出,这两类细菌与真核细胞是由一个原始的生物分别起源的不同的种类。研究者已经抛弃了这个模型,但是三域系統获得了普遍的认同。这样,细菌就可以被分为几个界,而在其他体系中被认为是一个界。它们通常被认为是一个单源的群体,但是这种方法仍有争议。
結構
繁殖
细菌以无性方式进行繁殖,最主要的方式是以二分裂法这种无性繁殖的方式:一个细菌细胞细胞壁横向分裂,形成两个子代细胞,在分裂的时候可以产生遗传重组。单个细胞也会通过如下几种方式发生遗传变异:突变(细胞自身的遗传密码发生随机改变),轉化(无修饰的DNA从一个细菌转移到溶液中另一个细菌中,并成功整合到该细菌DNA或质粒上,使之具有新的特征),轉染(病毒的或细菌的DNA,或者两者的DNA,通过噬菌体这种载体转移到另一个细菌中),细菌接合(一个细菌的DNA通过两细菌间形成的特殊的蛋白质结构,接合菌毛,转移到另一个细菌)。细菌可以通过这些方式获得基因片段,通过分裂,将重组的基因组传给后代。许多细菌都含有异源的DNA片段。
当细菌处于温度、湿度、空气、营养等丰富的环境中时,会快速繁殖,呈指数级增长,可以形成肉眼可见的集合体,例如菌落(colony)。
有些细菌可以形成芽孢结构,芽孢能够耐受高温、干旱、强辐射等极端恶劣,有利于其度过严峻的环境,保持自身的延续。
代谢
细菌具有许多不同的代谢方式。一些细菌只需要二氧化碳作为它们的碳源,被称作自养或自營生物。那些通过光合作用从光中获取能量的,称为光合自养生物。那些依靠氧化化合物中获取能量的,称为化能自养生物。依靠有机物形式的碳作为碳源,称为异养或異營生物。
異營細菌需從體外攝取有機物以維生,若有機物為屍體,則稱為腐生菌;若其有機物來自活體,則稱為寄生菌。自營細菌可以利用光能或化學能,將無機物自行合成有機養分。
光合自养菌包括蓝细菌(蓝藻),它是已知的最古老的生物,可能在制造地球大气的氧气中起了重要作用。其他的光合细菌进行一些不制造氧气的过程。包括绿硫细菌,绿非硫细菌,紫细菌和大腸桿菌。
正常生长所需要的营养物质包括氮,硫,磷,维生素和金属元素,例如钠,钾,钙,镁,铁,锌和钴。
根据它们对氧气的反应,大部分细菌可以被分为以下三类:一些只能在氧气存在的情况下生长,称为需氧菌;相反,只能在缺氧气情况下生长的,称为厌氧菌;还有些无论有氧无氧都能生长,称为兼性厌氧菌。细菌也能在人类认为是极端的环境中旺盛得生长,这类生物被称为嗜极生物。例如生存在于温泉中,称为嗜热细菌;一些生存在高盐湖中,称为嗜盐生物;还有存在酸性或碱性环境中,称为嗜酸细菌和嗜碱细菌;更有存在于阿尔卑斯山冰川中,称为嗜冷细菌。
細菌分裂速度相當快,約每隔數十分鐘即分裂一次,故在短短一小時內,就有2-3代產生。由於驚人的增值速度,可能會造成其所需的養分不足,並且形成過多代謝廢物,會妨礙細菌的生長,甚至造成死亡。
运动
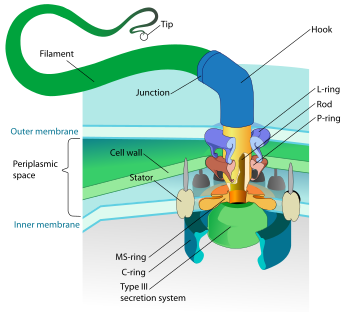
革兰氏阴性细菌的鞭毛。底座驱动钩和长丝的旋转。
运动型细菌可以依靠鞭毛,细菌滑行或改变浮力来四处移动。另一类细菌,螺旋體,具有一些类似鞭毛的结构,称为轴丝,连接周质的两细胞膜。当他们移动时,身体呈现扭曲的螺旋型。螺旋菌則不具軸絲,但其具有鞭毛。
细菌鞭毛以不同方式排布。细菌一端可以有单独的极鞭毛,或者一丛鞭毛。周毛菌表面具有分散的鞭毛。
运动型细菌可以被特定刺激吸引或驱逐,这个行为称作趋性,例如,趋化性,趋光性,趋磁性、趋机械性等。在一种特殊的细菌,粘细菌中,个体细菌互相吸引,聚集成团,形成子实体。
成長與繁殖
個體分裂
遺傳學
行為
细菌分类
细菌可以按照不同的方式分类。细菌具有不同的形状。大部分细菌是如下三类:杆菌是棒状;球菌是球形(例如链球菌或葡萄球菌);螺旋菌是螺旋形。另一类,弧菌,是逗号形。
细菌的结构十分简单,原核生物,没有膜结构的细胞器例如线粒体和叶绿体,但是有细胞壁。根据细胞壁的组成成分,细菌分为革兰氏阳性菌和革兰氏阴性菌。“革兰氏”来源于丹麥細菌學家漢斯·克里斯蒂安·革兰,他发明了革兰氏染色。
有些细菌细胞壁外有多糖形成的荚膜,形成了一层遮盖物或包膜。荚膜可以帮助细菌在干旱季节处于休眠状态,并能储存食物和处理废物。
與其他生物的交互關係
用處和危害
細菌對環境、人類和動物而言可說既有利又有害。一些細菌成為病原體,導致了破傷風、傷寒、肺炎、梅毒、食物中毒、霍亂,甚至是肺結核。在植物中,細菌導致葉斑病、火疫病和萎蔫。感染方式包括接觸、空氣傳播、食物、水和帶菌微生物。病原體可以用抗生素處理,抗生素分為殺菌型和抑菌型。但一般而言約百分之80%的細菌對人是無害的。
細菌通常與酵母菌及其他種類的真菌一起用於醱酵食物,例如在醋傳統的製造過程就是利用空氣中的醋酸菌(Acetobacter)使酒轉變成醋。其他利用細菌製造的食品還有乳酪、泡菜、醬油、醋、酒、酸奶等[28][29]。細菌也能夠分泌多種抗生素,例如鏈黴素即是由鏈黴菌(Steptomyces)所分泌的[30]。
細菌能降解多種有機化合物的能力也常被用來清除污染,稱做生物修復。舉例來說,科學家利用嗜甲烷菌來分解美國喬治亞州的三氯乙烯和四氯乙烯污染[30]。
历史
细菌这个名词最初由德国科学家埃倫伯格在1828年提出,原文「bacteria」一詞是新拉丁語,為「bacterium」的複數格。是由希臘文βακτήριον(bakterion)拉丁化而來[31]。意為“小棍子”[32],因為一開始發現的細菌是桿狀的。[33][34]
1866年,德国动物学家海克尔建议使用“原生生物”,包括所有单细胞生物(细菌、藻类、真菌和原生动物)。
1878年,法国外科医生塞迪悅(Charles Emmanuel Sedillot,1804-1883)提出“微生物”来描述细菌细胞或者更普遍的用来指微小生物体。
因为细菌是单细胞微生物,用肉眼无法看见,需要用显微镜来观察。1683年,列文虎克最先使用自己设计的单透镜显微镜观察到了细菌,大概放大200倍。路易斯·巴斯德和罗伯特·科赫指出细菌可导致疾病。
参见
- 細菌結構
- 細菌分類表
- 三域系統
- 古細菌
- 真核生物
- 細菌形態分類
參考文獻
引用
^ Bacteria (eubacteria). Taxonomy Browser, US National Institute of Health. [10 September 2008].
^ Whitman W, Coleman D, Wiebe W. Prokaryotes: the unseen majority. Proc Natl Acad Sci U S A. 1998, 95 (12): 6578–83. PMID 9618454.
^ Schulz H, Jorgensen B. Big bacteria. Annu Rev Microbiol: 105–37. PMID 11544351.
^ Sears C. A dynamic partnership: Celebrating our gut flora. Anaerobe. 2005, 11 (5): 247–51. PMID 16701579.
^ Fredrickson J, Zachara J, Balkwill D; 等. Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the hanford site, Washington state. Appl Environ Microbiol. 2004, 70 (7): 4230–41. PMID 15240306. 引文格式1维护:显式使用等标签 (link)
^ Cynthia Needham, Mahlon Hoagland, Keeneth McPherson, Bert Dodson. 觀念生物學3. 科學天地. 譯者:李千毅. 天下出版. 2005. ISBN 986-417-449-5.
^ http://science.nasa.gov/science-news/science-at-nasa/2007/11may_locad3/
^ Rappé M, Giovannoni S. The uncultured microbial majority. Annu Rev Microbiol: 369–94. PMID 14527284.
^ Whitman WB, Coleman DC, Wiebe WJ. Prokaryotes: the unseen majority. Proceedings of the National Academy of Sciences of the United States of America. 1998, 95 (12): 6578–83. Bibcode:1998PNAS...95.6578W. PMC 33863. PMID 9618454. doi:10.1073/pnas.95.12.6578.
^ C.Michael Hogan. 2010. Bacteria. Encyclopedia of Earth. eds. Sidney Draggan and C.J.Cleveland, National Council for Science and the Environment, Washington DC
^ 11.011.111.2 Choi, Charles Q. Microbes Thrive in Deepest Spot on Earth. LiveScience. 17 March 2013 [17 March 2013].
^ Glud R, Wenzhöfer F, Middelboe M, Oguri K, Turnewitsch R, Canfield DE, Kitazato H. High rates of microbial carbon turnover in sediments in the deepest oceanic trench on Earth. Nature Geoscience. 2013, 6 (4): 284. Bibcode:2013NatGe...6..284G. doi:10.1038/ngeo1773.
^ Oskin, Becky. Intraterrestrials: Life Thrives in Ocean Floor. LiveScience. 14 March 2013 [17 March 2013].
^ Schopf JW. Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic. Proceedings of the National Academy of Sciences of the United States of America. 1994, 91 (15): 6735–42. Bibcode:1994PNAS...91.6735S. PMC 44277. PMID 8041691. doi:10.1073/pnas.91.15.6735.
^ DeLong EF, Pace NR. Environmental diversity of bacteria and archaea. Syst Biol. 2001, 50 (4): 470–8. PMID 12116647. doi:10.1080/106351501750435040.
^ Brown JR, Doolittle WF. Archaea and the prokaryote-to-eukaryote transition. Microbiology and Molecular Biology Reviews. 1997, 61 (4): 456–502. PMC 232621. PMID 9409149.
^ Poole AM, Penny D. Evaluating hypotheses for the origin of eukaryotes. BioEssays. 2007, 29 (1): 74–84. PMID 17187354. doi:10.1002/bies.20516.
^ Dyall SD, Brown MT, Johnson PJ. Ancient invasions: from endosymbionts to organelles. Science. 2004, 304 (5668): 253–7. Bibcode:2004Sci...304..253D. PMID 15073369. doi:10.1126/science.1094884.
^ Lang BF, Gray MW, Burger G. Mitochondrial genome evolution and the origin of eukaryotes. Annu Rev Genet. 1999, 33: 351–97. PMID 10690412. doi:10.1146/annurev.genet.33.1.351.
^ McFadden GI. Endosymbiosis and evolution of the plant cell. Current Opinion in Plant Biology. 1999, 2 (6): 513–9. PMID 10607659. doi:10.1016/S1369-5266(99)00025-4.
^ Schulz HN, Jorgensen BB. Big bacteria. Annu Rev Microbiol. 2001, 55: 105–37. PMID 11544351. doi:10.1146/annurev.micro.55.1.105.
^ Williams, Caroline. Who are you calling simple?. New Scientist. 2011, 211 (2821): 38–41. doi:10.1016/S0262-4079(11)61709-0.
^ Robertson J, Gomersall M, Gill P. Mycoplasma hominis: growth, reproduction, and isolation of small viable cells. J Bacteriol. 1975, 124 (2): 1007–18. PMC 235991. PMID 1102522.
^ Velimirov, B. Nanobacteria, Ultramicrobacteria and Starvation Forms: A Search for the Smallest Metabolizing Bacterium. Microbes and Environments. 2001, 16 (2): 67–77. doi:10.1264/jsme2.2001.67.
^ Dusenbery, David B. (2009). Living at Micro Scale, pp. 20–25. Harvard University Press, Cambridge, Mass. ISBN 978-0-674-03116-6.
^ Fritz I, Strömpl C, Abraham WR. Phylogenetic relationships of the genera Stella, Labrys and Angulomicrobium within the 'Alphaproteobacteria' and description of Angulomicrobium amanitiforme sp. nov. Int J Syst Evol Microbiol. 2004, 54 (Pt 3): 651–7. PMID 15143003. doi:10.1099/ijs.0.02746-0.
^ Wanger G, Onstott TC, Southam G. Stars of the terrestrial deep subsurface: A novel 'star-shaped' bacterial morphotype from a South African platinum mine. Geobiology. 2008, 6 (3): 325–30. PMID 18498531. doi:10.1111/j.1472-4669.2008.00163.x.
^ Johnson M, Lucey J. Major technological advances and trends in cheese. J Dairy Sci. 2006, 89 (4): 1174–8. PMID 16537950.
^ Hagedorn S, Kaphammer B. Microbial biocatalysis in the generation of flavor and fragrance chemicals. Annu Rev Microbiol: 773–800. PMID 7826026.
^ 30.030.1 Cynthia Needham, Mahlon Hoagland, Keeneth McPherson, Bert Dodson. 觀念生物學4. 科學天地. 譯者:李千毅. 天下出版. 2005. ISBN 986-417-450-9.
^ βακτήριον. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project.
^ βακτηρία in Liddell and Scott.
^ bacterium, on Oxford Dictionaries.
^ Harper, Douglas. bacteria. Online Etymology Dictionary.
书籍
- Alcamo, I. Edward. Fundamentals of Microbiology. 5th ed. Menlo Park, California: Benjamin Cumming, 1997.
- Atlas, Ronald M. Principles of Microbiology. St. Louis, Missouri: Mosby, 1995.
- Holt, John.G. Bergey's Manual of Determinative Bacteriology. 9th ed. Baltimore, Maryland: Williams and Wilkins, 1994.
- Stanier, R.Y., J. L. Ingraham, M. L. Wheelis, and P. R. Painter. General Microbiology. 5th ed. Upper Saddle River, New Jersey: Prentice Hall, 1986.
- Witzany, G. Bio-Communication of Bacteria and their Evolutionary Roots in Natural Genome Editing Competences of Viruses. Open Evolution Journal 2: 44-54., 2008.
外部連結
维基物种中的分类信息:细菌 |
从维基百科的姊妹计划 了解更多有关 “细菌”的内容 | |
| 维基词典上的字词解释 | |
| 维基共享资源上的多媒体资源 | |
| 维基新闻上的新闻 | |
| 维基语录上的名言 | |
| 维基文库上的原始文献 | |
| 维基教科书上的教科书和手册 | |
| 维基学院上的學習资源 | |
MicrobeWiki, an extensive wiki about 細菌和病毒
- Bacteria that affect crops and other plants
- Bacterial Nomenclature Up-To-Date from DSMZ
Genera of the domain Bacteria – list of Prokaryotic names with Standing in Nomenclature- 最大的細菌
- Tree of Life: Eubacteria
- 細菌游泳和翻滾的影片
細菌星球 史蒂芬·古爾德著- 微生物線上教材
- Animated guide to bacterial cell structure.
- Bacteria Make Major Evolutionary Shift in the Lab
- Online collaboration for bacterial taxonomy.
PATRIC, a Bioinformatics Resource Center for bacterial pathogens, funded by NIAID
Bacterial Chemotaxis Interactive Simulator – A web-app that uses several simple algorithms to simulate bacterial chemotaxis.
Cell-Cell Communication in Bacteria on-line lecture by Bonnie Bassler, and TED: Discovering bacteria's amazing communication system
Sulfur-cycling fossil bacteria from the 1.8-Ga Duck Creek Formation provide promising evidence of evolution's null hypothesis, Proceedings of the National Academy of Sciences of the United States of America. Summarized in: Scientists discover bacteria that haven't evolved in more than 2 billion years, LiveScience and BusinessInsider
| 關於細菌 的圖書館資源 |
|
| ||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||

